
Введение
Недавнее появление на черном рынке наркотических веществ, так называемых новых психоактивных веществ (НПВ) – соединений, которые по различным причинам никогда не были выведены на фармацевтический рынок – становится все большей проблемой для регуляторных органов и клинических и судебно-медицинских токсикологических учреждений. Фактически, не подпадая под уголовное преследование по факту производства, торговли и употребления наркотических веществ в связи с отсутствием правовых актов, регулирующих производство, торговлю и распространение данных веществ, а также высокая скорость изменения структур соединений, распространение НПВ на черном рынке достигло больших объемов. Хотя новейшие аналитические методики, основанные на масс-спектрометрии высокого разрешения, высокоэффективной жидкостной хроматографии и ядерном магнитной резонансе могут значительно сократить время, необходимое для идентификации и характеристики данных молекул, было проведено сравнительно небольшое количество исследований, посвященных характеризации потенциально психоактивных соединений.
Наиболее распространёнными организмами для изучения эффектов химических субстанций являются грызуны. Иными словами, модели, основанные на использовании грызунов, широко использовались для изучения фармакотоксикологических эффектов НПВ, поскольку данные модели демонстрируют широкий спектр хорошо узнаваемых поведенческих реакций и позволяют оценивать различные физиологические и неврологические параметры [6, c. 3685-3709; 20, c. 132-144; 22, c. 174-188; 41, c. 130; 33, c. 68-82]. Более того, для грызунов характерно различное пространственное поведение, например шаблонное поведение и пространственные предпочтения, а также грызуны демонстрируют сложные социальные взаимодействия [3, c. 239-256]. Однако широкое использование грызунов ограничено высокой стоимостью проведения экспериментов, сложностью обращения с грызунами, большим количеством времени проведения экспериментов, а также этическими проблемами, возникающими в результате целенаправленного тестирования заведомо нетерапевтических соединений на млекопитающих, что, как ожидается, вызовут дискомфорт и страдания у животных.
В последнее десятилетие ученые из разных стран сообщают об использовании рыбок данио в качестве новой модели исследований в области нейропсихофармакологии как дополнительного подхода помимо млекопитающих [17, c. 1925-1944]. Данио рерио могут предоставить исследователям такие важные преимущества, как быстрое внеутробное развитие эмбрионов, большое количество эмбрионов (от десятков до сотен эмбрионов от одной пары) за один период оплодотворения [18, c. 253-310], малый размер, невысокая стоимость содержания и высокий уровень сходства с млекопитающими [14, c. 63-75].
В частности, мальки зебрафиш используются для скрининга психоактивных веществ, так как через 4 дня после оплодотворения у мальков развиваются все группы нейрональных клеток и их проекции, а на 5 день после оплодотворения у мальков можно исследовать локомоторную активность, так как они проявляют функциональную анатомическую и сенсомоторную активность с возможностью свободного движения [8, c. 85-111; 24, c. 525-541; 29, c. 3]. В отличие от мышей, данио рерио в основном активны в дневное время [7, c. 1-5] и изменения освещенности вызывают у них ряд стереотипных двигательный реакций, объединяемых под общим термином «визуальный двигательный ответ», и эти реакции используются исследователями для характеристики поведенческих реакций на ксенобиотики и лекарственные средства [4, c. 2526-2539; 11, c. 792-813]. Известно, что психостимулирующие препараты D-амфетамин и метамфетамин вызывают аналогичные локомоторные нарушения как у рыбок данио, так и у грызунов, а именно вызывают двухфазный дозозависимый эффект [12, c. 84-90; 21, c. 97-104]. Кроме того, подобно моделям грызунов, мальки зебрафиш использовались для изучения поведенческих реакций как ответ на введение каннабиса [19, c. 1-10].
Психические расстройства являются наиболее частыми эффектами употребления НПВ - синтетических каннабиноидов. В частности, в данной работе исследовались эффекты 5F-APINAC (5F-AKB-57, адамантан-1-ил 1-(5-фторпентил)-1Н-индазол-3-карбоксилат).
5F-APINAC является НПВ, проявляющим каннабимиметическую активность. На данный момент данных по токсичности, вызываемой данным веществом, нет, но на основании его структуры, схожей с синтетическими каннабиноидами (присутствие группы индазола-тетраметилциклопропилкетона), эффектами могут быть галлюцинации, возбуждение, сонливость, рвота и тахикардия. Недавнее исследование фармакодинамики 5F-APINAC in vivo показало, что он метаболизируется по тому же пути, что и PB-22 [27, c. 1193-1202; 28, c. 359-368].
Доклинические исследования 5F-APINAC на модели мальков зебрафиш могут быть полезны в области судебной медицины для оценки возможных эффектов данных веществ и проведения токсикологических исследований в случаях интоксикации НПВ.
Главной целью данной научной работы было исследование возможности использования мальков зебрафиш как быструю, простую и высокопроизводительную поведенческую модель для первичного скрининга НПВ. С этой целью, мы изучали ответ на введение 5F-APINAC у мальков зебрафиш.
Материалы и методы
Все экспериментальные процедуры проводились в соответствии с государственными и международными регуляторными актами: ГОСТ 33219-2014 «Руководство по содержанию и уходу за лабораторными животными. Правила содержания и ухода за рыбами, амфибиями и рептилиями», «Европейская конвенция о защите позвоночных животных, используемых для экспериментов или в иных научных целях» CETS 123 и Council Directive 86/609/EEC of 24 November 1986 on the approximation of laws, regulations and administrative provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes.
Самцы и самки зебрафиш дикой линии AB (European Zebrafish Resourse Center, Германия) содержались в отделе судебной медицины Университета Вероны. Рыбы содержались в аквариумах с постоянным обменом воды. Максимальная плотность: 1 особь на 2 литра воды. Рыбы содержались при световом цикле 14 часов:12 часов (день:ночь). Температура воды и воздуха поддерживались на постоянном уровне (28±0.5°С и 25°С, соответственно). Рыб кормили дважды в день сухим кормом (TetraMin, Tetra GmbH, Германия) и трижды в неделю живым кормом (Аква Меню, Россия).
Приготовление растворов
Мальки зебрафиш подвергались воздействию широкого диапазона доз (0.001-10 мкМ). Изначально, вещества растворяли в диметилсульфоксиде (ДМСО), и далее, приводили к рабочим концентрациям раствором для содержания мальков Е3 (конечная концентрация ДМСО – 0,1%). Контрольные группы подвергались воздействию только раствора-носителя (0,1% ДМСО в Е3). Концентрации веществ были выбраны согласно предыдущим исследованиям [15, c. 70-80]. Мальки зебрафиш пятидневного возраста помещали в 96-луночный микропланшет (1 особь на лунку, каждая лунка была заполнена 600 мкл среды Е3). Введение веществ проводили на шестой день после оплодотворения в 11 часов утра. Временные рамки введения веществ были выбраны таким образом, чтобы исключить влияние утреннего включения света на двигательную активность [7, c. 2-5]. Также сообщается, что голодание вплоть до 10 дня после оплодотворения не влияет на рост и выживаемость мальков [10, c. 515-518].
Поведенческие тесты
Локомоторная активность мальков зебрафиш регистрировалась в течение 3 дней с пятых до седьмых суток после оплодотворения. В первый день регистрировались базовые линии суточной активности. После введения препарата, регистрировали двигательную активность в течение следующих 42 часов. Данный позволил определить: а) острый ответ на введение вещества (активность проявляется в первые 4 часа после введения); б) долгосрочное влияние на суточный ритм двигательной активности.
Спонтанную активность регистрировали на видео со скоростью 25 кадров в секунду при постоянной подсветке инфракрасным светом с использованием высокопроизводительной системы видеотрекинга (DanioVision, Noldus, Нидерланды). Планшет с мальками помещали в камеру системы видеотрекинга, в которой поддерживали температуру 28°С с помощью термостата (Noldus, Нидерланды). Был установлен 12-ти часовой световой период (свет включался в 6:00, выключался в 18:00). В качестве источника света использовали матрицу из белых светодиодных ламп; интенсивность освещения – 0.17 Вт/м2. Чтобы минимизировать влияние отсутствия некоторых кадров, был установлен порог чувствительность 0.2 мм/кадр. Активность каждой зебрафиш рассчитывалась как общее расстояние, пройденное за 10 минут.
Анализ выживаемости
Оценка жизнеспособности мальков зебрафиш проводилась определением любой смертности, спровоцированной введением веществ. Сразу же после поведенческого анализа, мальков переносили в чашки Петри, заполненные средой для эмбрионов. Показатели выживаемости в каждой группе определялись ежедневно вплоть до 11-дневного возраста, то есть спустя пять дней после введения веществ.
Статистический анализ
Эффект 5F-APINAC на визуальный двигательный ответ оценивали с помощью дисперсионного анализа (ANOVA), принимая общее пройденное расстояние в качестве зависимой переменной и дозу вещества в качестве независимой переменной. Для сравнения эффектов различных доз использовали тест множественных сравнений Тьюки.
Результаты
Кратковременное воздействие 5F-APINAC (0.001-10 мкМ) на мальков зебрафиш дозозависимо уменьшало общее пройденное расстояние (табл.). Низкие дозы (0.001, 0.01, 0.1, 1 мкМ) вызывали снижение локомоторной активности при остром воздействии (рис. 1), в то время как при длительном воздействии (рис. 2), тренд кривых повторяет шаблон группы контроля, однако различия в пройденном расстоянии были обнаружены статистическими методиками (табл., рис. 3). Также, воздействие самой высокой дозой (10 мкМ) вызывало резкое снижение подвижности сразу после введения препарата (рис. 1), которое сохранялось вплоть до второго дня воздействия препарата (рис. 2).
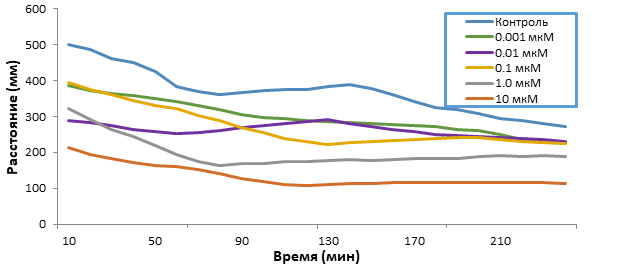
Рис. 1. Значения (среднее ± стандартное отклонение) пройденных расстояний мальками зебрафиш во время 4-х часового воздействия, выраженных в миллиметрах
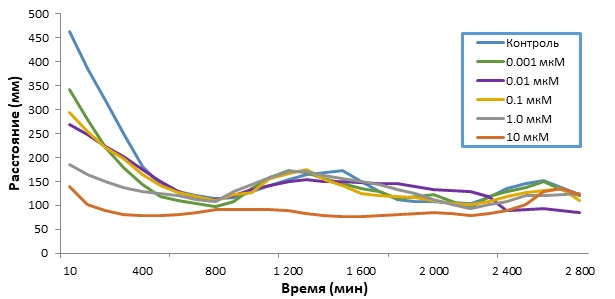
Рис. 2. Значения (среднее ± стандартное отклонение) пройденных расстояний мальками зебрафиш во время 48-ми часового воздействия, выраженных в миллиметрах
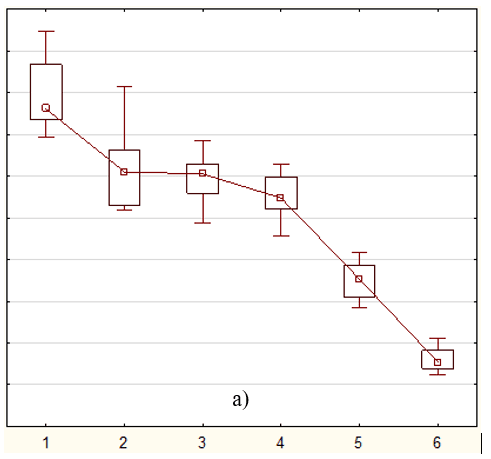
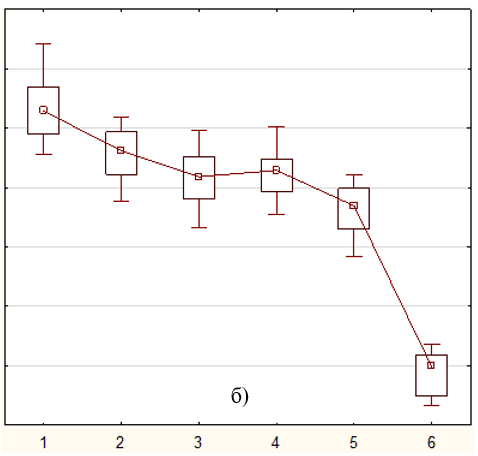
Рис. 3. Значения (среднее ± стандартная ошибка) общих пройденных расстояний мальками зебрафиш во время: а) 4-х часового воздействия; б) 48-ми часового воздействия. Обозначения: 1 – контроль, 2 – 0,001 мкМ, 3 – 0,01 мкМ, 4 – 0,1 мкМ, 5 – 1,0 мкМ, 6 – 10 мкМ.
Анализ набора данных после длительного воздействия показал, что на активность мальков зебрафиш влияло время дня: в дополнение к результатам 4-х часовой модели, эта модель показала, что животные были более активны в течение дневного времени, что и следовало ожидать от этого вида. Интересно, что самая высокая доза ухудшала способность синхронизировать двигательную активность с дневной фазой, что проявлялось в отсутствии разницы в пройденном расстоянии в темную и светлую фазы (рис. 2).
Таблица
Значения (среднее ± стандартное отклонение) общих пройденных расстояний мальками зебрафиш во время 4-х часового воздействия и 48-ми часового воздействия – статистический анализ.
|
Общее пройденное расстояние | ||
|
Группа воздействия |
Среднее ± стандартное отклонение |
p уровень значимости по сравнению с контролем |
|
4-х часовое воздействие | ||
|
Контроль |
9957,3 ± 885,5 |
– |
|
0,001 мкМ |
8140,2±882,7 |
НЗ* |
|
0,01 мкМ |
7969,8±530,4 |
НЗ* |
|
0,1 мкМ |
7518,0±533,5 |
>0,01 |
|
1,0 мкМ |
5496,1±461,3 |
>0,001 |
|
10 мкМ |
3608,4±285,8 |
>0,001 |
|
48-ми часовое воздействие | ||
|
Контроль |
51718,4±3018,6 |
— |
|
0,001 мкМ |
47797,4±2271,9 |
НЗ* |
|
0,01 мкМ |
45747,2±2515,5 |
>0,01 |
|
0,1 мкМ |
46283,6±2156,1 |
>0,05 |
|
1,0 мкМ |
43150,3±2098,0 |
>0,001 |
|
10 мкМ |
29406,8±1783,8 |
>0,001 |
*НЗ – статистически не значимо
Смертность более 50% была выявлена на девятый день после оплодотворения после введения высоких доз 5F-APINAC (1 и 10 мкМ). Морфологических изменений у выживших зебрафиш обнаружено не было.
Обсуждение
В данной работе мы исследовали поведенческие изменения, вызываемые новым психоактивным веществом 5F-APINAC, являющимся синтетическим каннабиноидом, у модели зебрафиш.
Бифазный эффект на двигательную активность (стимуляция при низких концентрациях и подавление при высоких), был описан у мальков данио. В зависимости от продолжительности воздействия и дозировки, агонисты каннабиноидных рецепторов модулируют спонтанную двигательную активность в виде двухфазного ответа, то есть с увеличением и уменьшением активности в низких и высоких дозировках, соответственно. Этот вид двухфазного ответа на введение препарата наблюдался при введении других агонистов каннабиноидных рецепторов: анандамида [32, c. 347-352], ∆9-ТГК [22, c. 174-188; 2, c. 283-293], WIN 55,212-2 [9, c. 145-150], JWH-018 [22, c. 174-188; 23, c. 130], AKB48, 5F-AKB48 [6, c. 3685-3709], 5F-ADBINACA, AB-FUBINACA и STS-135 [5, c. 1-15], и предполагает, что данный вид ответа типичен для каннабиноидной системы, а не для самих молекул [25, c. 483-501]. Двигательные нарушения и акинезия являются одними из наиболее известных поведенческих эффектов, наблюдаемых после введения каннабимиметиков [25, c. 483-501], причем эти эффекты в основном обусловлены стимуляцией каннабиноидных рецепторов первого типа мозжечка и базальных ганглиев [26, c. 703-713]. Поведенческая стимуляция, вызываемая синтетическими каннабиноидами, может быть связана с их способностью к увеличению высвобождения дофамина (ДА) в прилежащем ядре, возможно, через стимуляцию мезоаккумбального дофаминергического пути через каннабиноидно-рецепторный механизм [23, c. 130]. В отличие от вышеупомянутых исследований, в данном эксперименте введение 5F-APINAC не вызывало бифазного ответа. Это различие может быть связано с использованными концентрациями вещества, или же с различиями в протоколах эксперимента (например, выбор времени суток, использованный штамм данио, интенсивность света).
У зебрафиш, введение 5F-APINAC дозозависимо уменьшало общее пройденное расстояние в течение первых 4-х часов после введения. Эти результаты согласуются с предыдущими исследованиями влияния ∆9-тетрагидроканнабинола (∆9-ТГК) на зебрафиш [1, c. 349-360]. Интересно отметить, что как ∆9-ТГК, так и каннабидиол (КБД), оказывают ингибирующий эффект на двигательную активность у взрослых зебрафиш, что проявлялось снижением скорости плавания и уменьшением пройденного расстояния после воздействия данных соединений [13, c. 1-8; 31, c. 109-119]. Наличие данных эффектов принято считать проявлением тревожного поведения. Однако недавние исследования показали, что воздействие каннабиноидов на мальков данио изменяло синаптическую активность в нервно-мышечных соединениях, снижало частоту сердечных сокращений, а также уменьшало силу реакции на звук [16, c. 10518]. Эти результаты могут указывать на то, что снижение активности мальков зебрафиш может достигаться не только из-за тревожного эффекта.
В нашем исследовании наибольшая использованная доза 5F-APINAC (10 мкМ) влияла на циркадный ритм мальков зебрафиш. Дальнейшие исследования циркадных ритмов у рыбок данио крайне важны для понимания того, может ли этот вид быть полезным для изучения изменений циркадного поведения, вызываемых каннабиноидами.
Также, мы исследовали уровень смертности от введения 5F-APINAC. Высокие концентрации 5F-APINAC при введении малькам зебрафиш вызывали заметную смертность на пятый день после обработки веществом. Причины смерти зебрафиш не исследовались нами, так как главной целью нашей работы было проведение поведенческого анализа.
Заключение
Острое воздействие 5F-APINAC дозозависимо снижает двигательную активность, что согласуется с литературными данными, полученными в экспериментах на грызунах. Следовательно, результаты исследования говорят о том, что данио рерио можно использовать как инструмент быстрого предварительного скрининга поведенческой и неврологической активности НПВ. Однако стоит отметить, что у каждой биологической модели есть свои сильные и слабые стороны. Обычно существует отрицательная корреляция между эволюционным родством с людьми и экспериментальными соответствиями.
Кроме того, следует учитывать тот факт, что результаты поведенческих анализов варьируются у рыбок данио [30, c. 160451], но среди доступных высокопроизводительных систем, обладание высокой пропускной способностью может смягчить эффекты межиндивидуальной изменчивости.
