.png&w=384&q=75)
Введение
Одним из необходимых условий для развития отдельных способностей и творческих возможностей человека является эмоциональная культура, которая определяет и желания, и намерения, и коммуникативные качества в любом виде деятельности. Любимая музыка, любимые мелодии являются наиболее ярким отражением эмоциональной культуры каждого индивидуума.
Для изучения индивидуальных эмоциональных потребностей впервые в мире в Лаборатории хаоса создан музыкальный классификатор эмоций EMUSE [1], позволяющий детально рассматривать влияния мелодий на гармонию восприятия и подбирать музыкальные мелодии в соответствии с запросом.
Звуковой сигнал благодаря многочисленным исследованиям [2, с. 446] признан оптимальным носителем эмоций в существующей пространственно-временной реальности. Восприятие человеком звуковой гармонии музыкальных произведений может сопровождаться разнообразными эмоциональными паттернами – полимодальными образами, аффектами и другими реакциями. В настоящей работе стоит задача исследовать отражение в активности мозга возникновения этих паттернов, чтобы понять природу эмоций, источники их происхождения, принципы организации нейронных сетей в процессах осознания эмоций и, главное, возможное влияние эмоций на развитие отдельных свойств сознания и творческих способностей.
Цель исследования
Ранее [3] авторами было показано, что эмоциональное восприятие музыкальных мелодий сопровождается одновременным развитием возбуждения высокочастотных бета- и гамма-ритмов ЭЭГ в сенсорной и в лимбической системах в ответ на звуковые эмоциональные сигналы. Это означает, что в таких условиях звуковой сигнал при восприятии сразу принимает эмоциональную окраску, которая может осознаваться только с участием более медленных дельта- и тета-ритмов ЭЭГ.
К настоящему времени, в основном, изучены особенности развития дельта- и тета-активности в системах когнитивного восприятия [4]. В научных публикациях также представлены данные о тета-активности в передних отделах коры головного мозга при эмоциональных переживаниях как положительной, так и отрицательной валентности [5, с. 181]. В клинических исследованиях предложена методика оценки развития депрессий по тета-ритму ЭЭГ [6, с. 81-89].
Ранее [3] авторы обратили также внимание на то, что именно любимая музыка в большей степени обеспечивает выход в подсознание по сравнению с мелодиями, вызывающими только эмоции положительной валентности. Поэтому авторы высказывают предположение, что во время прослушивания любимых музыкальных мелодий могут возникать особые психо-эмоциональные состояния, связанные с подсознанием и способствующие развитию творчества.
В связи с этим, перед проведением настоящих исследований, посвящённых изменениям медленноволновой мозговой активности во время прослушивания любимых музыкальных мелодий, поставлены следующие задачи:
- Как изменяется спектральная мощность дельта- и тета-ритмов ЭЭГ и в каких областях коры мозга;
- Как музыкальный классификатор эмоций детально раскрывает эмоциональное содержание прослушиваемой музыки;
- Какие структуры лимбической системы возбуждаются при усилении дельта- и тета-активности в коре мозга;
- Какие структуры системы восходящей неспецифической активации возбуждаются при усилении дельта- и тета-активности в коре мозга;
- Какие структуры объединяются в системы взаимодействия нейронных сетей по ведущей частоте?
Материалы и методы исследования
В исследовании участвовала группа добровольных испытуемых, занимающихся интеллектуальным трудом (18 чел., из них 9 женщин и 9 мужчин), в возрасте от 30 до 60 лет. Регистрация электроэнцефалограммы (ЭЭГ) проводилась в состоянии психической и мышечной релаксации с закрытыми глазами и во время прослушивания в наушниках с закрытыми глазами любимой музыкальной мелодии, индивидуальной для каждого испытуемого.
Для регистрации ЭЭГ использовали 24-канальный нейровизор NVX24 производства ООО «Медицинские компьютерные системы» с применением стандартных монополярных отведений в соответствии с принятой международной схемой 10–20 [7, с. 376-382]. Исследования проводились в стандартных условиях, одинаковых для всех испытуемых.
Система обработки данных
Для обработки данных регистрации ЭЭГ использовали пакеты компьютерных программ, обеспечивающих спектральный анализ регистрируемых сигналов и топографическое картирование спектральных характеристик ЭЭГ [8, с. 65]. В результате получали распределение показателей мощности частотного спектра по отдельным областям на поверхности полушарий мозга. Статистический анализ данных проводится для определения достоверности полученных как индивидуальных, так и групповых усреднённых результатов.
С целью выявления наиболее выраженных реакций вычисляли локализацию и распределение эквивалентных дипольных источников (ЭДИ) происхождения электрической активности в коре и в глубинных структурах мозга. В некоторых работах этот процесс обозначен как эквивалентный токовый диполь (ЭТД). Для определения в объёме мозга человека локализации центров электрической активности программа вычисляла ЭДИ в каждый момент времени, равный дискретному интервалу отсчёта данных при вводе в компьютер, затем уровень активности оценивался по коэффициенту дипольности [9, с. 70]. Вопрос о соответствии локализации источников в глубинных структурах мозга той биоэлектрической активности, которая регистрировалась на поверхности головы, решался специальным алгоритмом, в котором рассматривались электрические свойства мозга как объёмного проводника [10, 11]. Для анализа применяли однодипольную модель ЭДИ. При интерпретации результатов работы программы учитывали, что вычисляемый для анализируемых одномоментных сечений ЭЭГ токовый диполь является эквивалентным, результирующим источником электрически активной в данный момент времени области мозга. ЭДИ характеризует суммарную электрическую активность нервных клеток, вовлеченных в текущий момент времени в процесс возбуждения, и расположен в электрическом центре этой области. Поэтому коэффициент дипольности, как показатель того, насколько точно вычисленная эквивалентная дипольная модель соответствует пространственному распределению потенциалов на поверхности головы, был использован на уровне высоких значений КД=>0,97. При этом ЭДИ потенциалов с таким высоким коэффициентом дипольности могут рассматриваться как указатели на наиболее активные локальные зоны, запускающие последовательное перемещение волновых процессов. Активные зоны в корковых структурах мозга обозначаются полями Бродмана [12]. Кроме того, в статье используются обозначения ядерных и ганглиозных структур мозга в составе лимбической системы и в составе неспецифической ретикулярной системы восходящей активации коры головного мозга.
Результаты исследования и их обсуждение
Для определения типа эмоционального насыщения музыкальных мелодий использован музыкальный классификатор эмоций EMUSE [1].
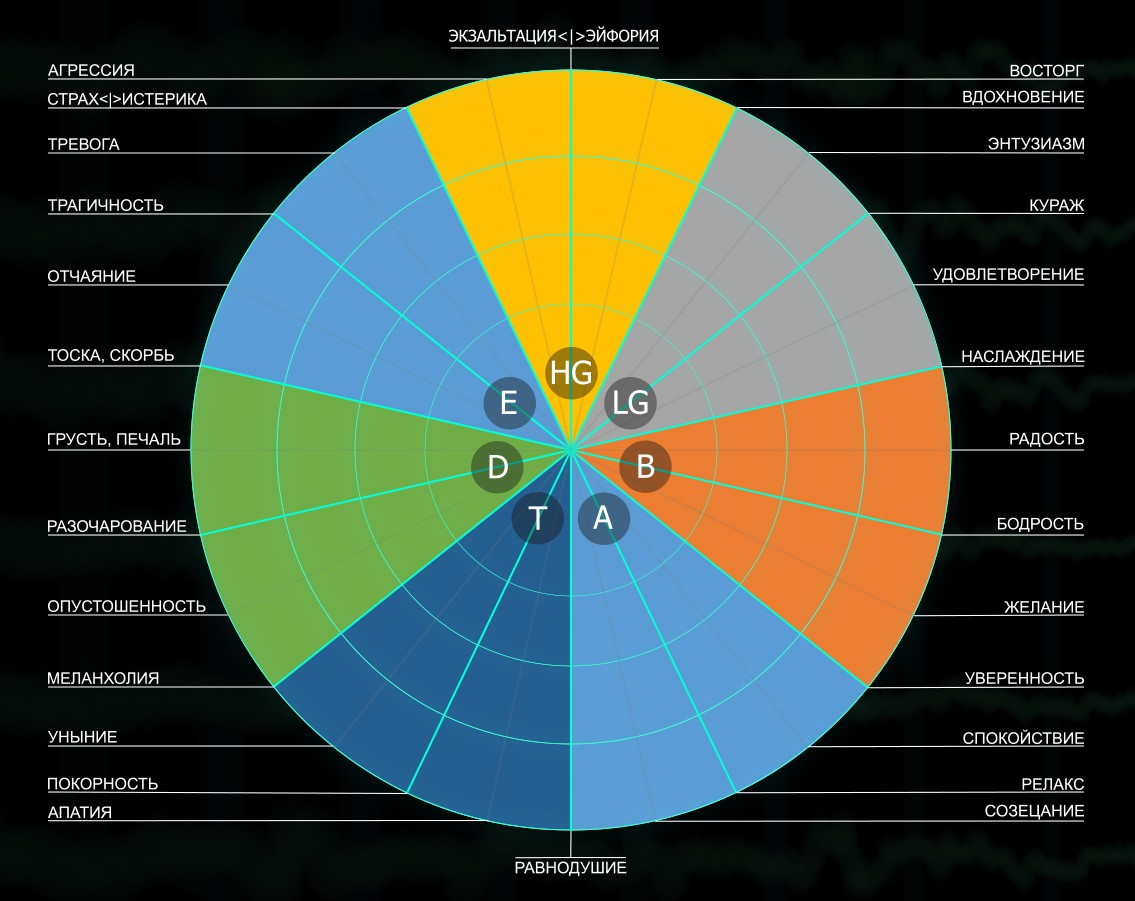
Рис. 1. Музыкальный классификатор эмоций EMUSE. Распределение эмоций на круговой шкале в соответствии с ритмами, характерными для активности мозга, по секторам: E – эпсилон 1-2 Гц; D – дельта 2–4 Гц; T – тета 4–8 Гц; A – альфа 8–16 Гц; B – бета 16–32 Гц; LG – низкая гамма 32–64 Гц; HG – высокая гамма 64–128 Гц
Круговая шкала эмоций на рисунке 1. поддерживается неспецифической реакцией AROUSAL (пробуждение) ретикулярной (пирамидной) и лимбической (экстрапирамидной) систем мозга. Радиальные шкалы внутри круговой шкалы определяют степень экспрессии эмоций, которые обозначены на круговой шкале. Внешняя окружность – это граница, внутри которой обозначены управляемые и саморегулируемые эмоции. За пределами окружности – патология или неуправляемые состояния и эмоции.
Музыкальные мелодии, особенно любимые, всегда многогранны и полимодальны. В данной работе обращаем внимание на диапазон медленных дельта- и тета-колебаний, которым на круговой шкале музыкального классификатора отведены сектора D и Т. В условиях прослушивания и усвоения музыкальных ритмов этим секторам соответствуют дельта- и тета-ритмы ЭЭГ.
У человека в норме концентрация медленноволновой активности наблюдается обычно в лобных и в передневисочных отделах коры головного мозга.
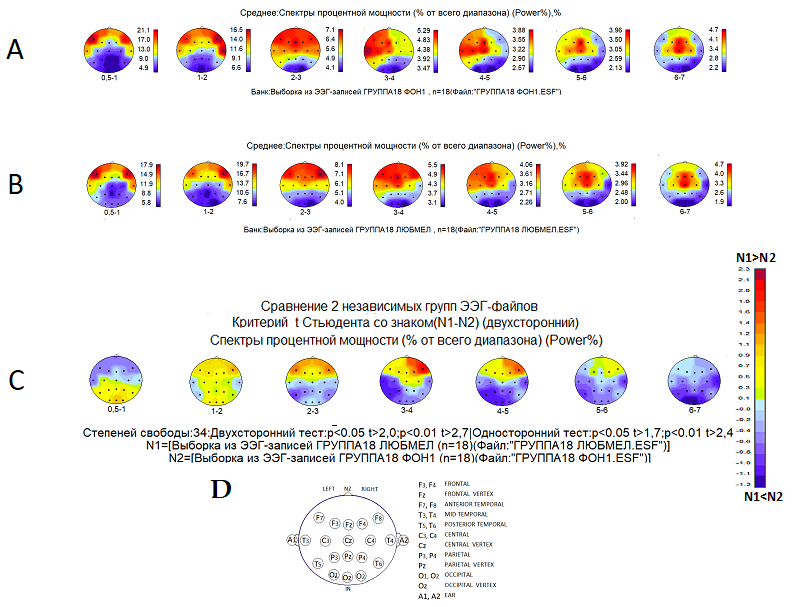
Рис. 2. Распределение спектральной мощности дельта- и тета-ритмов ЭЭГ в коре головного мозга (усреднённые данные группы испытуемых): А – в фоновом состоянии психической и мышечной релаксации в наушниках с закрытыми глазами; В – в состоянии прослушивания любимой мелодии в наушниках с закрытыми глазами; С – результаты статистического сравнения усреднённой спектральной мощности дельта- и тета-активности ЭЭГ: отличия в состоянии В по отношению к состоянию А; D – схема расположения электродов для регистрации ЭЭГ на поверхности головы
У испытуемых, участвующих в данном исследовании, в фоновом состоянии спокойного бодрствования, психической и мышечной релаксации с закрытыми глазами усреднённые данные показывают, что распределение спектральной мощности дельта-ритма ЭЭГ имеет максимумы на поверхности головы в обоих полушариях (рис. 2. А) в лобно-височных зонах частотой 0,5-1-2-3 Гц. Максимальные показатели спектральной мощности тета-ритма частотой 3-4 Гц и 4-5 Гц превалируют в лобной, височной и центральной областях левого полушария, а частотой 5-6 Гц и 6-7 Гц в зонах межполушария Fz и Cz. Таким образом, полученные результаты фоновой активности ЭЭГ группы испытуемых не противоречат норме.
В той же группе наблюдается аналогичное распределение спектральной мощности медленноволновой активности ЭЭГ при прослушивании любимых мелодий (рис. 2. В). Ранее в исследованиях [13, с. 288-297] было отмечено, что обычно у людей активность основных нейронных сетей мозга, наблюдаемая в фоновом состоянии, является устойчивой и мало изменяется при выполнении когнитивных задач или других ментальных действий, характерных для повседневной жизни. В определённой степени это относится и к прослушиванию музыкальных мелодий. Однако при относительно устойчивом распределении спектральной мощности дельта- и тета-ритмов в настоящем исследовании обнаружены статистически достоверные изменения процентного отношения ко всему диапазону спектральной мощности отдельных ритмов (рис. 2. С). В связи с этим, возникло предположение, что произошли изменения в организации распределения медленноволновой активности и в её распространении по нейронным сетям во время эмоционального прослушивания любимой музыки.
В целом, как показано на рисунке 2., формирование субъективного компонента эмоциональной реакции сопровождается избирательным вовлечением отдельных поддиапазонов дельта- и тета-активности мозга. Представленные достоверные изменения спектральной мощности наблюдаются в диапазонах частот дельта (2-3 Гц, 3-4 Гц) и тета-ритмов (4-5 Гц) в передних областях коры преимущественно правого полушария. Известно, что для правого полушария характерно целостное восприятие образов любой модальности. Установлено также [14, с. 41-54], что музыка влияет на оба полушария, при этом правое преимущественно воспринимает тембр и мелодию, а левое - ритмы.
Чтобы определить, с чем связаны изменения активности именно тех дельта- и тета-ритмов, которые обусловили различия между фоновым состоянием и состоянием при прослушивании любимой музыки (рис. 1. С), проведены исследования локализации эквивалентных дипольных источников (ЭДИ) их происхождения. Для этого были выделены ЭДИ с высоким коэффициентом дипольности (КД=>0,98), обозначающие локализацию наиболее активных центров возбуждения (рис. 3).

Рис. 3. Испытуемый Т. Пример распределения ЭДИ в нейроструктурах мозга в фоновом состоянии психической и мышечной релаксации (слева) и во время прослушивания любимой музыкальной мелодии (справа). Обозначения: ВА – поля Бродмана, HS – полушарие головного мозга, RF – ретикулярная формация, DEX – правый, SIN – левый. Изображения мозга в декартовой системе координат
Представленный на рисунке 3 пример отражает нарастающие во время прослушивания любимой музыкальной мелодии процессы возбуждения в полях Бродмана новой, старой и древней коры головного мозга, а также отдельных ядер таламуса, ретикулярной формации и особенно лимбической системы, включая базальные ганглии. Данное наблюдение с очевидностью показывает, что, прежде всего, активируется система эмоционального возбуждения и регулирующие функции передних областей коры головного мозга. Такая картина наблюдается практически у всех испытуемых.
Однако и сама любимая музыка, и индивидуальный подход к её восприятию у испытуемых разные. В силу вступает закон вероятности. Это означает, что у каждого испытуемого вероятность активации той или иной нейроструктуры индивидуальна и может отличаться при прослушивании музыкальной мелодии от фона. Из общего множества активных нейроструктур, на локализацию которых указывали ЭДИ с КД=>0,98, были выделены зоны с увеличенной вероятностью повышения активности во время звучания музыкальной мелодии по сравнению с фоновым состоянием психической и мышечной релаксации.
К этим зонам относятся поля Бродмана (ВА-Brodman area) ВА:11,18,25,28,34,36 и ядерные образования: хвостатое ядро (Nucleus Caudatus), миндалевидное тело (Corpus Amygdaloideum), чечевицеобразное ядро (Nucleus Lentiformis), гиппокамп (hippocampus), гипоталамус (hypothalamus), прилежащее ядро (nucleus accumbens). Перечисленные структуры относятся к лимбической системе, одной из основных функций которой является генерация эмоций. Ведущую роль в возбуждении и регуляции эмоций играет неокортекс [15, с. 8508-8513].
В таблице 1 представлены те нейроструктуры, которые были выделены на основании увеличенной вероятности активного возбуждения во время звучания мелодии по сравнению с фоном. Для сравнения указаны проценты участия данных активных нейронных структур в обоих состояниях испытуемых. В большинстве сопоставлений (в 28 из 35) имеется процентное преобладание случаев повышенной активности нейронных структур при прослушивании музыкальных мелодий, при этом звёздочками отмечены наиболее выраженные отличия (более чем на 60%, т. е. чаще, чем в два раза).
Таблица 1
Процентное соотношение участия структур мозга в активации нейронных сетей в фоне и при прослушивании любимых мелодий в группе испытуемых
Названия структур | Частота ритмов ЭЭГ | |||||
Коры головного мозга | 2-3 Гц | 3-4 Гц | 4-5 Гц | |||
Лимбической системы | Фон | Любмел | Фон | Любмел | Фон | Любмел |
ВА 11 | 54% | 46% | 45% | 55% | 31% | 69% * |
ВА 18 | 13% | 87% * | 31% | 69% * | 61% | 39% |
ВА 25 | 37% | 63% * | 39% | 61% * | 64% | 36% |
ВА 28 | 44% | 56% | 11% | 89% * | 43% | 57% |
ВА 34 | 30% | 70% * | 29% | 71% * | 40% | 60% |
ВА 36 | 58% | 42% | 44% | 56% | 35% | 65% * |
Nucleus caudatus | 35% | 65% * | 45% | 55% | 39% | 61% * |
Corpus amygdaloideum | 29% | 71% * | 20% | 80% * | 33% | 67% * |
Nucleus lentiformis | 34% | 66% * | 21% | 79% * | 54% | 46% |
Hippocampus | 60% | 40% | 25% | 75% * | 33% | 67% * |
Hypothalamus |
| 100% * |
| 100% * |
| 100% * |
Nucleus accumbens | 20% | 80% * |
|
| 50% | 50% |
Средний процент | 33% | 66% | 26% | 66% | 40% | 60% |
Если рассмотреть реакцию лимбической системы на отдельных частотах дельта- и тета-ритмов при прослушивании любимой музыкальной мелодии, то можно отметить, что в диапазоне дельта-ритма частотой 2-3 Гц повышается вероятность участия хвостатого ядра, миндалевидного комплекса, чечевицеобразного ядра, гипоталамуса и прилежащего ядра, а также возрастает вероятность участия в запуске функциональной активности полей Бродмана 18, 25, 34; на частоте 3-4 Гц дельта-ритма в запуске активных нейросетей участвуют миндалевидный комплекс, чечевицеобразное ядро, гипоталамус, гиппокамп и поля Бродмана 18, 25, 28, 34. В диапазоне тета-ритма 4-5 Гц во время прослушивания музыки по сравнению с фоном наиболее активное влияние на нейросети оказывают хвостатое ядро, миндалевидный комплекс, гиппокамп, гипоталамус и поля Бродмана 11, 36.
Таким образом, групповой анализ данных распределения ЭДИ дельта- и тета-ритмов показывает участие наиболее активных центров возбуждения в лимбической системе и в коре головного мозга в запуске последующих нейросетей, обеспечивающих интуитивное и эмоциональное восприятие.
При этом дельта- и тета-активность в образованиях ретикулярной системы продолговатого мозга (Medulla oblongata), Варо́лиева моста (Varolius), среднего мозга (Mesencephalon) и таламуса (Thalamus), включая субталамическое ядро (Nucleus subthalamicus), сохраняется при прослушивании любимой музыки, в основном, на том же уровне, как и в фоне. Однако во время эмоциональных переживаний возрастает вероятность повышенного возбуждения красного ядра (Nucleus Ruber) в три раза на частоте 2-3 Гц и чёрной субстанции (Substantia Nigra) в два раза на частоте 3-4 Гц, по сравнению с фоном. Эти два ретикулярных образования имеют нисходящие и восходящие анатомические связи непосредственно с миндалевидным комплексом и другими базальными ганглиями и обеспечивают их контакты с образованиями спинного мозга и таламуса. Ядра передней, передневентральной и переднемедиальной областей таламуса служат для прямой и обратной передачи сигналов в префронтальную и фронтальную кору от хвостатого ядра, близрасположенных структур и базальных ганглиев. Таким образом, анатомические и функциональные связи, прежде всего, лимбической системы с передней корой головного мозга, создают условия для эмоционального сопровождения восприятия любимой музыкальной мелодии.
Что касается индивидуального характера восприятия каждого испытуемого, входящего в группу, то можно говорить о развитии процесса эмоционального сопровождения с определённой долей вероятности, опираясь на данные таблицы 1.
Однако с более высокой точностью можно проследить индивидуальное развитие реакций мозга на эмоциональное прослушивание музыкальной мелодии с помощью построения пути перехода во времени от одного центра повышенной активности к следующему. Эта точность достигает интервала времени дискретизации ЭЭГ, которое осуществляется во время преобразования аналог-код для цифровизации и форматирования записи.
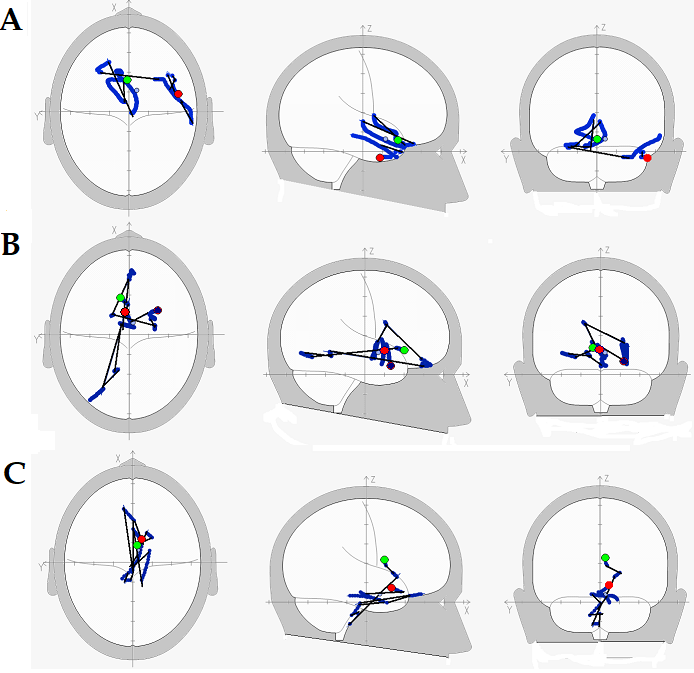
Рис. 4. Испытуемый Г. Последовательность максимального возбуждения центров нейронной активности во время прослушивания любимой музыкальной мелодии. Распределения ЭДИ при КД=>0,99. Зелёный ЭДИ – возбуждаемый в начале прослушивания, красный – в конце. ЭДИ происхождения ритмов ЭЭГ частотой: А –2-3 Гц; В – 3-4 Гц; С – 4-5 Гц. Путь А. S:BA 11,47, Putamen, Globus Pallidus lateralis, Globus Pallidus medialis, Hypothalamus, Nucleus ventralis anterior, Nucleus anterior, ВА 11,47,11, Gyrus Rectus, inter-hemispherum; D: Gyrus Frontalis medialis, Substantia Nigra, Nucleus Ruber, Nucleus dorsalis medialis; S:BA 47,38; D:BA 38,21,20,21,38,21. Путь В. S: BA 32,25,19,18,мозжечок, 19,11,Inter-Hemispherum; D: BA 11; S: Nucleus caudatus; D: Putamen, Corpus Amygdaloideum, Hippocampus, Putamen, Corpus Amygdaloideum, Putamen, BA 34,28; S: Nucleus ventralis anterior, Hypothalamus, Corpus Mamillaris, Nucleus ventralis anterior, Hypothalamus; D: Corpus Mamillaris; S: BA 25, Hypothalamus. Путь С. D: Nucleus caudatus, Putamen; S: мозжечок, Medulla oblongata, Varolius; BA11; D: Varolius, Mesencephalon, BA 36,35,28, Varolius, мозжечок, BA11,25. Обозначения: S – левое полушарие, D – правое полушарие, ВА – поля Бродмана
Наличие определенного уровня энергетического потенциала одной и той же частоты в разных звеньях сети проявляется в совпадении узкополосных частотных пиков на спектрограммах электрической активности корковых зон. В ранее опубликованной работе [16, с. 28-34] это явление обозначено как «энергетическая» составляющая пространственной организации биопотенциалов. При этом для взаимодействия активных нейронных центров и организации нейронных сетей имеет значение синхронизация их возбуждения по частоте.
Имеются наблюдения, что при восприятии невербальной информации ведущим мозговым механизмом выступает пространственная синхронизация мозга.
Существование медленных колебательных процессов обеспечивается тем, что сам процесс управляет поступлением необходимой для его поддержания энергии, и в этом смысле процесс является автоколебательным [17, с. 240].
Таблица 2
Повышенная активность нейроструктур в нейросетях при прослушивании любимых музыкальных мелодий
№ | 2-3Hz | 3-4Hz | 4-5Hz |
1 | ВА 18s-34d-NLs-AMd-Hipd | BA 11d-28s-NCd | BA 36d-HIPs |
2 | BA 11s-11d-25s-25d-NLs-NCs | BA 11s-11d-25s | BA 11s-34s |
3 | BA 11d-11s-25s-25d-36d-NCs-NCd-NLs-NLd | BA 11d-11s-18s-25s-25d-36d-NCd-NCs-AMd-NLd-NLs-HIPd | BA 11s-25d-36d-NCd-NLd |
4 | BA 11d-18d25d-28s-34s-34d-NCs-NCd-AMs-AMd-NLs-NLd | BA 28d-NCs-NLs | BA 11d-18s-25d-28s-28d-34d-36s-NCd-AMd-NLs-NLd-HYPs-HYPd |
5 | BA 18s-18d-25s-25d-36s-NCd-NLd-NACd | BA 18s | BA 18s-18d-36s-36d-NCs |
6 | BA 28d-34d-AMs | BA 11s-11d-18s-25s-25d-28d-34s-34d-36d-NCs-NCd-NLs-NLd-HIPs-HIPd-HYPd-HYPs | NCs-NCd-NLs-NLd |
7 | BA 11s | BA 11s-NCd | BA 11s |
8 |
|
| BA18s-34d-36d |
9 | BA NCd |
| BA 36d |
10 | NLd-NACd | BA 11s-11d-18s-18d-NCs | BA 18s-NLd-HIPd |
11 | BA 11s-11d-18d-25d-25s-34d-NCd-NLd-HIPd | BA 11s-18d-36d-NLs-NLd | BA11s-18d-NCd-NLd |
12 | BA 11s-25s-25d-28d-NCs-NLd-HYPd | BA 36s | BA 18s-34d-36d |
13 | BA36d | BA36s | NCd |
14 | BA18s-18d | BA18s-18d | BA18d-36s-Hips |
15 | BA 11s-25s-25d-28d-34d-36d-NCs-NCd-NLs-NLd | BA 28d-34d-36d-NCd-AMd-HIPd | BA 11d-NCd |
16 | BA 28d | BA 28s-AMs | BA 11d |
17 | BA 11d-11s-25s-25d-34s-36d-NCs-NCd-AMd-NLs-NLd-NACs | BA 11s-11d-25s-25d-28s-28d-34s-34d-36s-36d-NCs-NCd-AMs-AMd-NLs-NLd-HIPs-HIPd-HYPs-HYPd | BA11s-11d-25s-25d-28s-28d-34s-34d-36s-36d-NCs-AMs-NLd-HIPs-HYPd-HYPs |
Обозначения в таблице 2: ВА – поля Бродмана, AM – Corpus Amygdaloideum, HIP – Hippocampus, HYP – Hypothalamus, NAC – Nucleus accumbens, NC – Nucleus caudatus, NL – Nucleus lentiformis, s – левое полушарие, d – правое полушарие. ЛЮБМЕЛ – любимая музыкальная мелодия, № – номера испытуемых.
В настоящем исследовании проверка возможности возникновения дельта-колебаний частотой 2-3 Гц, 3-4 Гц и тета-колебаний частотой 4-5 Гц как следствия амплитудной или частотной модуляции ритмов высокочастотной активности мозга, дала отрицательный результат. Возможны нейрохимические и нейрогуморальные основы возникновения дельта- и тета-ритмов, учитывая состояние повышенной активности гипоталамуса.
В таблице 2 перечислены наиболее активные нейроструктуры каждого испытуемого во время прослушивания музыки. В таблице отражены индивидуальные комплексы нейроструктур, которые активируются на каждой частоте дельта- и тета-ритма относительно независимо. Каким образом они включаются в деятельность нейросетей?
По данным Н. Е. Свидерской [16, с. 28-34], даже при одинаковых узкополосных частотных характеристиках потенциалов в разных корковых зонах наличие «энергетического» фактора является необходимым, но недостаточным условием функционирования нейронных сетей. Когерентные взаимосвязи биопотенциалов как резонансные контуры не будут функционировать, если нет временной согласованности в работе его отдельных элементов. В таком случае прохождение сигнала блокируется, так как его приход может совпадать с периодом рефрактерности «принимающего» или «передаточного» звеньев. Это нарушит циркуляцию импульсов, и передача информации окажется невозможной. Таким образом, вторым условием для функционирования резонансных систем является временная, т. е. фазовая настройка электрических колебаний.
Однако мозг – живая самоадаптирующаяся система. Широко известны в нейрофизиологии реакции усвоения ритмов разнообразных внешних сигналов (световых, звуковых и др.) в рамках частоты естественных ритмов мозга. Аналогичные процессы «усвоения ритмов» могут происходить во время взаимодействия между нейроструктурами. Например, сам эффект альфа-стробирования [18, с. 33-39] выполняется за счёт сонастройки во времени активности нейроструктуры при прохождении альфа-волны сенсорного сигнала для придания ему необходимых свойств.
В более общем варианте этот процесс можно представить следующим образом. На частоте дельта- и тета-ритмов образуются объёмные электрические волны, которые регистрируются с поверхности головы в виде ЭЭГ. Параметры этих волн используются для вычисления эквивалентных дипольных источников их происхождения (ЭДИ) в глубинных структурах мозга. Чем больше по величине параметр, чем ближе к вершине волны он измеряется, тем более высокий коэффициент дипольности ЭДИ и тем более высокий уровень активности в глубинных структурах он отражает. Максимальная вершина поверхностной волны и её ЭДИ – это одномоментные явления. Наблюдения показывают, что вершинам дельта- или тета-волн обычно соответствуют несколько ЭДИ, следующих друг за другом, этим они создают условия для резонансного захвата. Поэтому можно допустить, что если у испытуемого перечисленные в таблице 2 нейроструктуры последовательно достигают максимального возбуждения на частоте дельта-ритма 2-3 Гц, то с высокой вероятностью они все продолжают работать в том же ритме, и резонансные эффекты на данной частоте возможны с каждой из перечисленных структур.
В другом варианте отдельные звенья нейросетей могут работать независимо друг от друга.
В более ранних исследованиях [19] были показаны принципиальные различия в активности мозга и организации взаимодействия слуховой зоны коры с другими областями у музыкантов и немузыкантов. По мнению авторов, это свидетельствует о специализации и относительной независимости осцилляторных систем мозга, участвующих в формировании различных компонентов эмоциональной реакции.
В отдельных исследованиях [20, с. 3-19] отмечено, что в ответ на понравившуюся музыку в мозге активируется несколько зон, но самым чувствительным оказалось прилежащее ядро (nucleus accumbens) – область, которая возбуждается, когда что-либо оправдывает наши ожидания. Прилежащее ядро связано и с другими областями мозга, а в случае со звуками задействована еще и слуховая кора. И чем больше нам нравятся те звуки, которые мы слышим, тем сильнее это взаимодействие, тем больше образуется новых нейронных связей, которые, как известно, и составляют основу наших когнитивных способностей. Но чтобы прогнозировать, какую именно мелодию предпочтет каждый конкретный человек, необходимо знать его музыкальные вкусы, за которые отвечает височная доля. Поэтому важно изучить связи и функциональные отношения между ней и прилежащим ядром. Не исключено, что некоторые функции прилежащего ядра осуществляются на подсознательном уровне.
Хвостатое ядро (nucleus caudatus) также является одной из структур мозга, которые составляют систему вознаграждения [20, с. 3-19]. Хвостатое ядро имеет отношение и к формированию эмоций, в том числе положительных, и способствует формированию привязанности.
Бледный шар в составе чечевицеобразного ядря (nucleus lentiformis) считается более древним образованием (палеостриатум), чем хвостатое ядро (неостриатум). Неостриатум реализует пусковой механизм поведения человека, т. е. осуществляет выбор действия и его программы, так как в процессе онтогенеза он совместно со скорлупой апробирует эти программы и накапливает их в своем арсенале. Он также регулирует вегетативное обеспечение и эмоциональное сопровождение двигательной активности.
Передние области мозга тесно связаны с головкой хвостатого ядра. Отмечается, что патология, возникающая в одной из взаимосвязанных областей системы кора – хвостатое ядро, функционально компенсируется сохранившейся структурой.
Роль лимбико-таламо-кортикального тета-ритма состоит в избирательной обработке значимой информации и одновременной защите этой обработки от интерференции, что можно интерпретировать как процессы селекции мишени внимания [21, с. 91-108].
Контроль следа внимания осуществляется со стороны фронтальной коры. Приведены данные [22, с. 560], подтверждающие существование в реакции внимания позднего компонента медленноволновой негативности, имеющего фронтальную или фронтоцентральную локализацию. Этот комронент может отражать ярко выраженное вовлечение фронтальной коры в процесс селективного внимания. Показана более высокая активность в правой фронтальной коре по сравнению с левой, что доказывает доминантность правого полушария в контроле функций внимания.
В ряде работ модуляции внимания связываются с осцилляциями на частоте тета-ритма 4-8 Гц (покорность, апатия, равнодушие) как этап системогенеза, сопровождающийся увеличением вклада низкодифференцированных систем в осуществление нового поведения.
Выводы:
- Формирование субъективного компонента эмоциональной реакции при прослушивании любимой мелодии сопровождается избирательным вовлечением отдельных поддиапазонов дельта- и тета-ритмов ЭЭГ в передних областях коры мозга преимущественно правого полушария. Это подтверждается достоверным увеличением спектра процентной мощности (% от всего диапазона) дельта-ритма (2-3 Гц, 3-4 Гц) и тета-ритма (4-5 Гц).
- В структурах лимбической системы выделены ведущие центры возбуждения нейросетей, обеспечивающих интуитивное и эмоциональное восприятие. Особое внимание в работе направлено на центры с увеличенной вероятностью повышения дельта- и тета-активности во время звучания музыкальной мелодии по сравнению с фоном. Показано, что при прослушивании любимой музыкальной мелодии повышается вероятность участия хвостатого ядра (nucleus caudatus), чечевицеобразного ядра (nucleus lentiformis), миндалевидного комплекса (corpus amygdaloideum), гиппокампа (hippocamp), гипоталамуса (hypothalamus) и прилежащего ядра (nucleus accumbens), а также возрастает вероятность участия в запуске функциональной активности полей Бродмана 11, 18, 25, 28, 34, 36. При этом индивидуальные различия эмоциональных реакций выражаются вероятностными соотношениями участия в них отдельных из перечисленных выше структур. Таким образом, большинство структур лимбической системы и отдельные области коры головного мозга имеют более высокую активность в дельта- и тета-диапазонах частот при прослушивании любимой музыкальной мелодии по сравнению с фоновым состоянием психической и мышечной релаксации.
- Дельта- и тета-активность в образованиях ретикулярной системы продолговатого мозга (Medulla oblongata), Варо́лиева моста (Varolius), среднего мозга (Mesencephalon) и таламуса (Thalamus), включая субталамическое ядро (Nucleus subthalamicus), сохраняется при прослушивании любимой музыки, в основном, на том же уровне, как и в фоне. Однако во время эмоциональных переживаний возрастает вероятность повышенного возбуждения красного ядра (Nucleus Ruber) в три раза на частоте 2-3 Гц и чёрной субстанции (Substantia Nigra) в два раза на частоте 3-4 Гц, по сравнению с фоном.
