.png&w=384&q=75)
Введение
Существуют разные способы зрительного восприятия без участия глаз. Некоторые из них имеют общие механизмы реализации.
Ранее было показано, что одним из вариантов зрительного восприятия без участия глаз могут быть синестезии [1, с. 91-95; 2, с. 19-29]. Были раскрыты биофизические особенности активности мозга при возникновении синестезий у лиц с доминирующим типом восприятия [2, с. 19-29; 3, с. 6-12]. Синестезии в этих работах рассматривались как способность преобразования сигналов, поступающих в мозг по сенсорному каналу одной модальности, в образы другой сенсорной модальности. Авторами также раскрыт механизм такого преобразования путём переноса сенсорной информации с участием альфа-сканирования и альфа-стробирования между каналами прохождения и первичными центрами восприятия сенсорных сигналов разных модальностей.
Появление современных технологий развития у людей видения без глаз и их практическое применение подтверждают, что зрительное восприятие может возникать без прямого использования глаз, но с обязательным участием зрительной системы мозга [4]. Например, сенсорное замещение (sensory substitution) было использовано при разработке устройств типа BrainPort или vOICe, преобразующих визуальную информацию видеокамер в тактильные или звуковые сигналы [5, с. 155-159; 6, с. 87-109; 7; 8]. Мозг со временем учится интерпретировать эти сигналы как «зрение». Слепые люди, использующие эхолокацию с помощью щелчков языком (как Дэниел Киш), задействуют затылочную долю для навигации. ФМРТ подтверждает, что мозг перестраивается под новые «модальности».
Видение без глаз возможно, если под «зрением» понимать субъективное восприятие визуальной информации, создаваемое мозгом. Ключевое условие – сохранность зрительной коры и её способности обрабатывать сигналы, независимо от их источника (импланты, другие органы чувств или внутренняя активность мозга). Технологии типа кортикальных имплантов или сенсорного замещения уже сегодня позволяют слепым «видеть» в ограниченной форме [7, 8].
Наименее изученным и наиболее перспективным направлением для понимания и развития ЭСВ является исследование так называемого «альтернативного» способа видения без участия глаз. Опубликованных работ, посвящённых изучению этой способности, очень мало, но необходимо выделить экспериментальные исследования в Московском государственном университете им. М. В. Ломоносова проф. Ю. П. Пытьева и др. [9; 10; 11, с. 43-45]. Эти работы показали, что альтернативное видение, как замена обычного, используется «в обход» прямого зрительного пути от глаз.
Необычное явление альтернативного видения у зрячих и у слабо видящих лиц до 2000 года воспринималось медицинским научным сообществом как невозможное и противоречащее основным постулатам биологии, нейрофизиологии и психологии. В связи с этим, под руководством академика РАН и РАМН Н. П. Бехтеревой была совершена попытка верифицировать сами факты альтернативного видения и проверить возможность измерения связанных с ними физиологических параметров [12, с. 43-49].
В исследованиях лаборатории Института мозга принимали участие 7 учащихся старших классов средней школы, прошедших обучение альтернативному видению по методу В. М. Бронникова. Все испытуемые продемонстрировали возможности альтернативного видения в различных условиях испытаний. Для обнаружения мозговых признаков феномена проводилось сравнение спонтанной электрической активности мозга (ЭЭГ) и вызванных потенциалов (ВП) во время выполнения испытуемыми однотипных заданий на классификацию предъявляемых изображений в состоянии обычного зрительного восприятия и в состоянии альтернативного видения.
Данные ЭЭГ подтвердили перестройку мозга на другой режим функционирования при проведении функциональных проб с «рассматриванием» предметов. Наличие сходных изменений ЭЭГ у разных лиц (при исходном различии их ЭЭГ) косвенно свидетельствовало о том, что речь идет не об уникальном феномене, а о воспроизводимом, обучаемом процессе. При исследовании ВП выявлены факты статистически достоверных различий ВП в условиях классификации изображений с открытыми глазами без маски и в маске на глазах. Например, у испытуемого В. Б. в начале исследований имела место, по данным ВП, более четкая дифференциация обычного зрительного и альтернативного видения. У него в начале исследований с высокой достоверностью проявлялись относительно коротколатентные ВП в затылочных областях при работе без маски, означающие поступление сигналов от глаз в зону первичного зрительного восприятия, но переставшие обнаруживаться при работе с маской [11, с. 43-45].
В результате проведённой работы не опровергли, а, наоборот, подтвердили наличие альтернативного видения у обученных испытуемых. Явление существует, оно воспроизводимо и может изучаться нейрофизиологическими методами. Можно также считать, что проведена апробация методик регистрации электрической активности мозга (ЭЭГ) и вызванных потенциалов (ВП) применительно к исследуемой проблеме.
Таким образом, нейрофизиологические исследования подтвердили существование феномена альтернативного видения. Показано, что «включение» альтернативного видения изменяет характер активности мозга.
Несколько ранее на кафедре компьютерных методов физики физического факультета МГУ им. М. В. Ломоносова было установлено, что изучаемый феномен альтернативного видения обладает свойствами волнового процесса и характеризуется двумя важными чертами, позволяющими для его исследования применять физические методы – полной воспроизводимостью и возможностью выполнять физические измерения [9, 10].
Эксперименты выявили голографический характер этого варианта ЭСВ и возможности его интерпретации в терминах, свойственных зрительной системе.
В работах проф. Ю. П. Пытьева и соавторов обсуждается «голографическая модель альтернативного видения, согласно которой испытуемые при ЭСВ порождают «опорное излучение», организованное как волновой процесс, и воспринимают (на полевом уровне) пространственное распределение интенсивности, возникающее при интерференции опорного и рассеянного объектами излучения, несущего «отпечаток» оптической картины последних. В данном случае ЭСВ рассматривается как активный (со стороны испытуемых) процесс, при котором интерпретация альтернативного видения осуществляется теми же функциями сознания, которые ответственны за обычное зрительного восприятия и создают перцептивный образ объектов, организуя их видение» [11, с. 43-45].
ЭСВ испытуемого В. Б. (школа В. М. Бронникова) и его зрительная интерпретация имеют определённые отличия от некоторых других видов ЭСВ. Он может воспринимать окружающий мир без использования глаз и интерпретировать восприятие в терминах зрительной системы практически адекватно обычному зрению.
Участвуя в экспериментах Ю. П. Пытьева, В. Б. воспринимает освещённые белым светом объекты с разрешением, достаточным для того, чтобы читать нормальный книжный печатный текст. Объекты воспринимаются им правильно окрашенными, то есть так же, как при обычном зрительном восприятии, их ориентация и размер не искажены, равно как и расстояние между ними.
Если предположить, что испытуемые с ЭСВ могут являться источником излучения опорной волны и обладать способностью воспринимать распределение интенсивности, возникающее в результате интерференции опорной и рассеянной объектами волн, то «видение объектов можно считать результатом интерпретации сознанием этого распределения интенсивности, т. е. голограммы.
В связи с гипотезой голографического восприятия [9; 10; 11, с. 43-45] проф. Ю. П. Пытьев и соавторы ставят следующие задачи исследования:
«Необходимо установить волновую природу рассматриваемого феномена экстрасенсорного восприятия, выяснить, где находится и как функционирует источник опорной волны, где и как воспринимается рассеянная волна, какова длина опорной волны, какова её природа, как рассеянная волна интерпретируется сознанием и т. д.».
Получены следующие результаты. В основе восприятия В. Б., как и в случае с другими испытуемыми, лежит волновой процесс. Опыты с дифракционной решеткой позволили определить длину волны порядка 1,5–2 мм в зависимости от состояния В. Б. [10]. Волновая природа феномена подтверждена также опытами с зонной пластинкой Френеля. Получаемые таким путём значения длины волны, полностью согласовывались с найденными ранее с помощью дифракционной решётки.
Для определения зон восприятия без участия глаз на поверхности тела было использовано экранирование, в результате которого выделялись области с ослабленным либо полностью прерванным «видением». У испытуемой Н. А. одной из основных областей оказалась центральная часть лба размером порядка 3–5 см по горизонтали, 8–4 см по вертикали. В несколько меньшей степени с «видением» оказались связаны области висков и макушки, области на груди. Вначале было предположение, что как излучение, так и восприятие могут быть связаны с областями на лбу, темени и на груди. Но поскольку маловероятно, что могут быть активными одновременно оба процесса в одном месте, то оставалось проверить, является ли область на лбу источником опорной волны.
Используя разные уровни изоляции предполагаемого опорного излучения из области лба от рассматриваемых без участия глаз объектов, авторам [9] удалось установить, что лобная область является у испытуемой Н. А. «только излучающей», следовательно, возможным претендентом на источник опорного излучения.
Однако у испытуемого В. Б. голографическое видение несколько отличается от Н. А. Что касается непосредственного восприятия объектных рассеянных волн и как эти волны интерпретируются сознанием, то с помощью специальных экспериментов было определено пространственное расположение искомых областей восприятия «анфас». «Глаза», определяющие «точку зрения», из которой В. Б. «видит», интерпретируя ЭСВ зрительно, обычно находятся непосредственно перед его глазами, на расстоянии порядка 15–20 см, если В. Б. без маски, и на расстоянии 25–30 см, когда В.Б. в маске. Но В.Б. может перемещать «виртуальные глаза» в пространстве, приближая их к тому или иному объекту и рассматривая его. Эта способность позволяет В. Б. «видеть» как бы сквозь непрозрачные предметы, помещая «глаза» за них.
В. Б. голографически (в целом) воспринимает объекты и среду, интерпретирует же восприятие зрительно, с той или иной «точки зрения», помещая в эту точку «виртуальные глаза».
Однако найденные области не следует буквально понимать как области восприятия. Речь идёт об областях, которые интерпретируются сознанием как «глаза» при стереовосприятии. Более того, как оказалось, любой предмет, помещённый в одну из найденных областей, «ослеплял» на один «глаз».
Впрочем, тот факт, что с закрытыми глазами В.Б. «видит» предметы окрашенными, как и при обычном видении, говорит о том, что в обоих случаях восприятие интерпретируется одними и теми же функциями сознания. Последние результаты исследований цветного зрения свидетельствуют о том, что интерпретация в терминах цвета, скорее всего, осуществляется на уровне коры и не определяется непосредственно цветочувствительными клетками сетчатки глаза.
По мнению проф. Ю. П. Пытьева, пока что наиболее ощутимым результатом этих работ стало обозначение феномена научным термином – экстрасенсорное восприятие. Иначе говоря, распознавание окружающего мира не с помощью пяти известных органов чувств, а посредством неких полей, окружающих человека.
На основе многочисленных результатов анализа активности мозга по данным ЭЭГ у лиц (детей-подростков и взрослых), прошедших специальную подготовку развития альтернативного видения по методу В. М. Бронникова, сделаны предварительные выводы о том, что происходит перестройка деятельности мозга, затрагивающая, прежде всего, фронтальные области коры и базальные отделы с центрами активации эмоций. Однако не все обучающиеся альтернативному видению достигают уровня чтения книги в маске, хотя могут уже рассмотреть отдельные крупные буквы или слова. Возникает предположение, что устойчивые показатели активности мозга у человека с наиболее устойчивой способностью читать книгу без использования глаз (более 30 лет у В. Б.) своего рода эталонные показатели, могут подсказать, в чём заключаются отличия у лиц, отстающих в обучении. Эти отличия позволят более чётко определить недостаточность развития отдельных функций и сконцентрировать на них внимание в процессе дальнейшего обучения. Контроль активности мозга является оптимальным подходом в текущей оценке развития альтернативного видения в процессе обучения.
Учитывая сложность и многофункциональность феномена альтернативного видения, авторы данной статьи решили продолжить изучение его проявления с наиболее стабильными результатами (около 30 лет) у испытуемого В. Б. и постараться найти пути к ответам на вопросы, поставленные ранее проф. Ю. П. Пытьевым и акад. Н. П. Бехтеревой:
- Не ясны механизмы генерации «опорного излучения» (генерируемого испытуемым) и его связь с сознанием. Волновая природа излучения остаётся загадкой, так как оно, возможно, сложнополяризованное, а, возможно, даже не электромагнитное, но взаимодействует с электромагнитными полями. При этом длина волны излучения зависит от состояния сознания испытуемого и влияет на эффективность альтернативного видения.
- Если видение внешних объектов связано с их взаимодействием с «виртуальными глазами», то пути поступления сигналов в центры зрительного восприятия мозга для запуска процесса создания осознаваемых образов остаются неизвестными. Предположение, что восприятие возникает за счёт интерференции «опорного излучения» (создаваемого испытуемым) с рассеянным светом от объектов – это возможное начало пути к осознанию альтернативного видения (но глаза при этом не видят). Тогда построение собственно зрительных образов в ответ на сигналы не от глаз может осуществляться с помощью синестезий [1, с. 91-95; 2, с. 19-29] или отдельных механизмов ассоциативной памяти.
Поэтому в данной статье сконцентрировано внимание на особенностях ЭЭГ-активности мозга испытуемого В. Б., подтвердившего свои способности в физических исследованиях проф. Ю. П. Пытьева и в нейрофизиологических исследованиях под руководством акад. РАН и РАМН Н. П. Бехтеревой. Возникла задача создания некоего «эталона» основных выделяющихся признаков деятельности мозга при осуществлении альтернативного видения. Сопоставление с этими «нормативными» признаками реальных показателей активности мозга у обучающихся лиц позволит выявить те функции мозга, к которым необходимо проявить дополнительное внимание для их развития.
В настоящей статье рассматривается выполнение следующих задач:
- Изучение особенностей биофизических характеристик отдельных структур мозга у испытуемого В. Б. с использованием методов вычисления эквивалентных дипольных источников (ЭДИ). Определение возможных признаков «опорного излучения» и механизмов его возникновения. Сопоставление активности центров сенсорного восприятия разных модальностей для выявления путей сенсорного замещения при альтернативном видении.
- Изучение спектральных характеристик ЭЭГ В. Б. Применение нейрофизиологического метода картирования данных на поверхности коры мозга для оценки изменения активности первичных центров сенсорного восприятия разных модальностей при альтернативном видении для выявления путей сенсорного замещения.
- сопоставление параметров, отражающих «эталонные» характеристики активности мозга в процессе альтернативного видения, с текущими параметрами мозговой активности у лиц, обучающихся альтернативному видению по методике В. М. Бронникова.
Методика
В качестве наиболее адекватных методов получения объективных данных о динамике состояний сознания выбраны нейрофизиологические способы регистрации активности головного мозга – электроэнцефалограммы (ЭЭГ). Известно, что классические описания таких состояний сознания человека, как сон и бодрствование, были получены по результатам исследований ЭЭГ [13, с. 23-24; 14, с. 1307-1321]. Поэтому для оценки особых состояний сознания, которые сопровождают экстрасенсорное восприятие в процессе альтернативного видения, также использовали ЭЭГ с дальнейшим применением компьютерных комплексов и программ по обработке сигналов мозга.
Биопотенциалы мозга регистрировали при закрытых глазах в состоянии психической и мышечной релаксации и в процессе чтения книги без использования глаз, закрытых светонепроницаемой маской.
Для оценки изменений ЭЭГ при тестовых испытаниях альтернативного видения использовали процедуру сравнения с ЭЭГ в фоновом состоянии психической и мышечной релаксации с закрытыми глазами. В этом случае фоновые показатели активности мозга становились началом отсчёта, так как условия регистрации ЭЭГ (освещение, уровень шумовых помех, температура воздуха, состояние организма и т. д.) оставались одинаковыми как в фоне, так и при выполнении тестов. Поэтому отличия в показателях, которые возникали при выполнении тестов, были отнесены непосредственно к тестируемым процессам. Но поскольку некоторая функциональная изменчивость ЭЭГ характерна и в фоновом состоянии, то обращали внимание только на статистически достоверные отличия.
Регистрацию ЭЭГ проводили на 24-канальном нейровизоре NVX24 производства ООО "Медицинские Компьютерные Системы" с использованием стандартных монополярных отведений в соответствии с принятой международной схемой 10–20% [15, с. 2-17].
Основная цель обработки данных регистрации ЭЭГ заключалась в сопоставлении самоотчёта испытуемого, наблюдаемых паттернов в ритмах мозга и локализации центров электрической активности, ответственных за происхождение этих паттернов. Для этого использовали пакеты компьютерных программ, обеспечивающих анализ спектральных характеристик ЭЭГ и топографическое картирование, а также трёхмерную локализацию источников электрической активности головного мозга [16, с. 376-382; 17]. Применяли статистический анализ данных для определения достоверности полученных изменений.
Для определения локализации источников электрической активности программа BL6 [17] вычисляла эквивалентные дипольные источники (ЭДИ) ритмической активности мозга по фильтрованным ЭЭГ-данным в дельта-, тета-, бета- и гамма-частотных диапазонах для каждого момента времени с шагом 0.002 сек. Достоверность результатов оценивали исходя из величины коэффициента дипольности (КД). Учитывались только ЭДИ с КД ≥ 0.94.
Результаты трёхмерной локализации ЭДИ графически обозначались точками на трёх ортогональных проекциях головы (вид сверху, вид сбоку и вид сзади) и выводились в текстовый листинг с указанием анатомической привязки ЭДИ к структурам головного мозга.
При интерпретации результатов работы программы имелось в виду, что вычисляемый для анализируемых одномоментных сечений ЭЭГ токовый диполь является эквивалентным, результирующим источником потенциалов ЭЭГ для электрически активной в данный момент времени области мозга. Эквивалентный диполь характеризует суммарную электрическую активность нервных клеток, вовлеченных в текущий момент времени в процесс электрического возбуждения, и расположен в электрическом центре этой области.
Коэффициент дипольности (Кд) является показателем того, насколько точно вычисленная дипольная модель описывает пространственное распределение потенциалов на поверхности головы в данный момент времени [17, 18]. Высокий уровень КД отражает не только точность и надёжность вычисления ЭДИ, но и повышенную суммарную электрическую активность в центре нейронного возбуждения. Поэтому при самых высоких значениях КД ≥ 0.98–0.99 наблюдаются только распределения ЭДИ электрических колебаний потенциала максимально активных нейроструктур мозга. С учётом того, что максимально активные нейронные структуры могут определять поведение нейросетей в текущий момент времени, этот показатель был использован для оценки и сравнения состояний сознания в процессе развития альтернативного видения.
Результаты и обсуждение
Нейрофизиологические исследования, проводимые с участниками школы развития альтернативного видения Вячеслава Михайловича Бронникова, показали определённые особенности развития активности мозга в процессе проявления феномена. Так как альтернативное зрение в настоящее время не является широко распространённой способностью среди людей, но обучение этому свойству сознания возможно, то особое значение приобретают признаки его проявления и особенности активности мозга при этом. Поэтому исследование было проведено на первом этапе с наиболее выраженной и устойчивой (более 30 лет) способностью к чтению книжного текста без участия глаз у испытуемого В. Б.
Регистрация ЭЭГ В. Б. проводилась в состоянии спокойного бодрствования с закрытыми глазами (фон) и в процессе чтения книги молча без участия глаз, закрытых светонепроницаемой маской, с последующим отчётом о содержания текста сразу после окончания регистрации ЭЭГ. Записи ЭЭГ подвергались не только спектральному анализу и оценке статистической достоверности изменений ЭЭГ во время чтения книги по сравнению с фоном, но прежде всего, было обращено внимание на изменения биофизических характеристик: распределение эквивалентных дипольных источников (ЭДИ) дельта-, тета- и гамма-ритмов ЭЭГ в коре головного мозга.
Рисунок 1 показывает, что у испытуемого В. Б. уже в фоновом состоянии психической и мышечной релаксации имеется повышенная локализация ЭДИ дельта- и тета-ритмов в лобно-височной области левого полушария, указывающая на концентрацию активности нейроструктур в этой области. Повышенная активность этой области в фоновом состоянии говорит о том, что она всегда находится в состоянии готовности. Во время чтения книги без использования глаз (в светонепроницаемой маске) активность в этой области значительно усиливается, что определяется по показателям увеличения процентного отношения количества ЭДИ при КД ≥ 0.94 к общему количеству вычисленных ЭДИ. При этом наблюдается увеличение по сравнению с фоном количества ЭДИ дельта-ритма (КД ≥ 0.94) с 9.6% до 12.0% и ЭДИ тета-ритма (КД ≥ 0.96) с 2.7% до 6.2%.
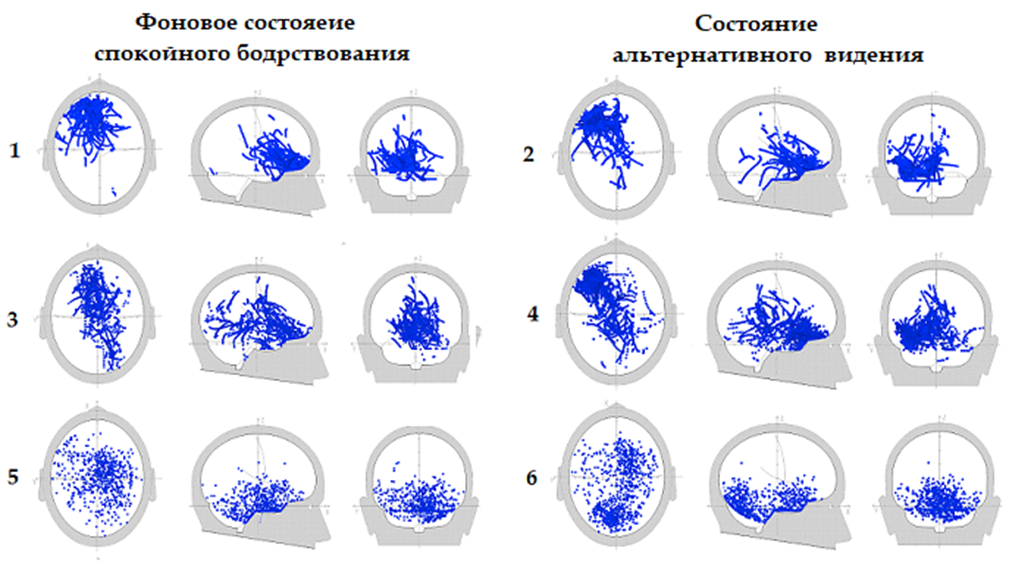
Рис. 1. Испытуемый В. Б. Распределения ЭДИ дельта, тета и гамма ритмов в структурах мозга, представленных в декартовой системе координат: 1) Фон – дельта 0.5–3.5 Гц, КД=0.94; 9.6%; 2) Внешнее видение – дельта 0.5–3.5 Гц, КД=0.94; 12.0%; 3) Ф он – тета 4–7 Гц, КД=0.96; 2.7%; 4) Внешнее видение – тета 4–7 Гц, КД=0.96; 6.2%; 5) Фон – гамма 30–45 Гц КД=0.96 1.2% 6) Внешнее видение – гамма 30–45 Гц, КД=0.96 1.6%
Необходимо отметить, что известны функциональные различия левого и правого полушарий мозга [18]. В целом, если правое полушарие доминирует в приёме поступающей информации в виде внешних сигналов в объёме гораздо большем, чем осознаётся, то в левом полушарии необходимая часть информации из правого подвергается анализу и формированию действия. Конечно, отдельные поля активности перекрываются по-разному у разных людей, но можно опираться на эти основные представления.
Более вероятно, что в левом полушарии формируются исходящие потоки активности, которые усиливаются при чтении книжного текста с непрозрачной маской на глазах (рис. 1 (1–4)). Необходимо дальнейшее рассмотрение значения этого параметра повышенной концентрации ЭДИ дельта- и тета-ритмов в левой лобно-височной области, как одного из возможных показателей альтернативного видения.
Иначе распределяются ЭДИ гамма-ритма. Если ЭДИ гамма-ритма, как показано на рисунке 1 (5-6), в фоновом состоянии сознания концентрируются преимущественно в центральной области правого полушария, то в состоянии альтернативного видения дополнительно усиливается их концентрация в затылочной области обоих полушарий.: количество выявленных ЭДИ при КД ≥ 0.96 увеличивается с 2.7% до 6.2%. Повышенная активность в правом полушарии как в фоне, так и во время чтения книги в маске говорит, скорее всего, о преобладании процессов обработки воспринимаемых сигналов, когда формируются зрительные образы книжного текста без участия глаз.
Таблица
Максимально активные структуры мозга за период регистрации ЭЭГ 100 с и количество моментов локализации ЭДИ с повышенным КД ≥ 0.98–0.99
Дельта-ритм в фоне | Дельта-ритм во время альтернативного видения | |||||||||
Левое полушарие | Правое полушарие | Левое полушарие | Правое полушарие | |||||||
ВА 11 | 89 |
|
| ВА 11 | 320 | ВА 11 | 15 | |||
|
|
|
| ВА 38 | 56 | ВА 32 | 13 | |||
|
|
|
| ВА 47 | 34 | ВА 25 | 15 | |||
Тета-ритм в фоне | Тета-ритм во время альтернативного видения | |||||||||
Левое полушарие | Правое полушарие | Левое полушарие | Правое полушарие | |||||||
ВА11 | 6 |
|
| ВА 11 | 55 |
|
| |||
ВА24 | 1 |
|
| ВА 38 | 23 |
|
| |||
|
|
|
| ВА 47 | 47 |
|
| |||
Гамма-ритм в фоне | Гамма-ритм во время альтернативного видения | |||||||||
Левое полушарие | Правое полушарие | Левое полушарие | Правое полушарие | |||||||
ВА11 | 2 | ВА11 | 2 | ВА 11 | 1 | ВА 18 | 1 | |||
ВА20 | 1 | ВА19 | 1 | ВА 18 | 1 | ВА 20 | 6 | |||
ВА22 | 1 | ВА20 | 3 | ВА 38 | 1 | ВА 21 | 1 | |||
ВА28 | 5 | ВА25 | 2 |
|
| ВА 28 | 1 | |||
ВА34 | 1 | ВА28 | 2 |
|
| ВА 36 | 4 | |||
ВА35 | 1 | ВА32 | 2 |
|
| ВА 38 | 9 | |||
ВА36 | 1 | ВА34 | 4 |
|
| ВА 47 | 3 | |||
ВА38 | 4 | ВА36 | 4 |
|
| Amygdalum | 1 | |||
ВА47 | 5 | ВА37 | 1 |
|
| Hypothalamus | 1 | |||
N. ruber | 1 | ВА47 | 1 |
|
|
|
| |||
|
| Amygdalum | 1 |
|
|
|
| |||
|
| C.mammilaris | 2 |
|
|
|
| |||
|
| Hypothalamus | 1 |
|
|
|
| |||
|
| N.lentiformis | 2 |
|
|
|
| |||
|
| Subs.nigrа | 1 |
|
|
|
| |||
В фоновом состоянии самым активным в диапазоне дельта-ритма ЭЭГ является поле Бродмана 11 (Brodman area – BA11) в левом полушарии. Эта часть орбитофронтальной коры, в отличие от других фронтальных областей, имеет обширные функциональные связи с другими областями коры, включая сенсорные системы восприятия разной модальности. Кроме того, BA11 участвует в принятии решений, функционировании системы вознаграждений, планировании, в переносе новой информации в долговременную память, в значительной степени способствует концентрации внимания. В таблице отмечено, что не наблюдается ни одной другой структуры, кроме BA11, ни в левом, ни в правом полушарии, которая достигла бы такого же уровня активности в фоновом дельта-ритме ЭЭГ. Надо отметить, что испытуемый В.Б. в обычной повседневной жизни использует как видение с помощью глаз, так и альтернативное видение без участия глаз. Возможно, поэтому характерная локализация ЭДИ дельта- и тета-ритмов в виде потоков, направленных к поверхности левого полушария, устойчиво сохраняется в обычном фоновом состоянии, значительно усиливаясь при альтернативном видении.
В состоянии альтернативного видения резко повышается активность BA11 в левом полушарии, достигая максимального уровня дипольности в течение 320 моментов за 100 с, по сравнению с 89 моментами в фоне. В левом полушарии проявляется также высокая активность BA38 и BA47. Известно, что BA38 участвует в семантическом представлении высокого уровня и социально-эмоциональной обработке данных.
Область Бродмана 38 (BA38), представляет собой структурно и функционально сложную область, расположенную в самой передней части височной доли. Часто называемая височно-полярной корой головного мозга, эта область играет решающую интегративную роль в обработке эмоций, семантической памяти и социальном познании. Благодаря богатой анатомической структуре и участию в различных когнитивных процессах височный полюс всё чаще рассматривается как зона конвергенции, связывающая потоки перцептивной, эмоциональной и концептуальной обработки информации. Существует прямой двунаправленный путь к орбитофронтальной коре, который позволяет мнемоническим представлениям, хранящимся в височном полюсе, влиять на принятие решений в лобной доле.
BA47 – часть лобной коры головного мозга, которая располагается латеральнее BA11 и изгибается вентрально. BA47 участвует в обработке синтаксиса в устной речи и семантических аспектах языка, а также в распознавании некоторых эмоций. Таким образом, можно предположить, что левое полушарие во время чтения книги с использованием альтернативного видения, с одной стороны, формирует внешний контакт с книжным текстом при участии BA11, с другой – проводит синтаксический и семантический анализ этого текста с помощью наиболее активных BA38 и BA47.
Дельта-ритм ЭЭГ отражает также активацию BA11, BA32, BA25 в правом полушарии. BA11 в правом полушарии значительно менее активен, чем BA11 в левом полушарии – почти в 21 раз. Этот факт подчеркивает мощную активацию BA11 в левой лобно-височной области и позволяет предположить возникновение особых свойств этой активации. Признаки максимальной активности дельта-ритма в BA32 – рецепторной области эмоций в поясной извилине, могут сопровождать эмоциональные проявления альтернативного видения, а максимум дельта-активности в BA25, которое является основным распределителем серотонина в мозге человека, и, следовательно, может стать участником сверхактивности в BA11 левого полушария.
Организация максимальной активности структур мозга в тета-диапазоне ритмов ЭЭГ, в основном, повторяет организацию в дельта-диапазоне ритмов. Отличия заключаются только в том, что в фоновом состоянии добавляется кратковременно возникшая максимальная активность BA24, а в состоянии альтернативного видения в правом полушарии не выявляются структуры мозга, которые достигли бы высокого уровня активности при максимальном КД.
Иная картина распределения структур с повышенной активностью наблюдается в диапазоне гамма-ритма ЭЭГ. Увеличивается общее количество высокоактивных структур с преобладанием в правом полушарии, как в фоновом состоянии, так и в состоянии альтернативного видения. По всей вероятности, это является следствием повышенной сенсорной деятельности обоих полушарий, особенно правого. В фоновом состоянии активны отдельные BA коры больших полушарий, поясной извилины и гиппокампа, отдельные ядерные образования лимбической системы. В целом, выделенная активность отражает равновесное состояние мозга во время спокойного бодрствования.
В состоянии альтернативного видения в правом полушарии преобладает активность тех структур, которые могут отражать внутреннее проговаривание читаемого текста (BA20, BA21), синтактический и семантический анализ текста (BA38, BA47), работу с памятью (BA28, BA36), пространственную ориентацию (BA18). Системы Amygdalum и Hypothalamus могут оказывать эмоциональную и гуморальную поддержку основным функциям. Левое полушарие менее активно.
В результате, в состоянии альтернативного видения создаются специальные условия в лобной области левого полушария – максимальная активность в BA11, где концентрируется активность дельта (2-3 Гц) и тета (4–7 Гц) ритмов ЭЭГ под влиянием ряда условий, в том числе нейрогуморальной активации со стороны BA25. Эти условия могут способствовать возникновению очага инфракрасного (ИК) излучения дальнего диапазона с длиной волны около 2 мм, установленной в лаборатории проф. Ю. П. Пытьева [9; 10; 11, с. 43-45]. Современные исследования подтверждают эффекты фотонного излучения мозга человека и значение в этом процессе митохондриальной активности [23, 24].
Показатели распределения спектральной мощности ритмов ЭЭГ (рис. 2) согласуются с данными локализации ЭДИ происхождения этих ритмов, представленными на рисунке 1.
На рисунке 2 представлены результаты статистического сравнения спектральных характеристик двух записей ЭЭГ испытуемого В.Б. в фоновом состоянии и во время альтернативного видения. Изменения в виде достоверного увеличения спектральной мощности ЭЭГ в диапазонах дельта- и тета-ритмов представлены в лобно-височной области левого полушария, где и наблюдается повышение плотности распределения ЭДИ этих ритмов во время чтения книги без глаз (рис. 1 (1–4)).
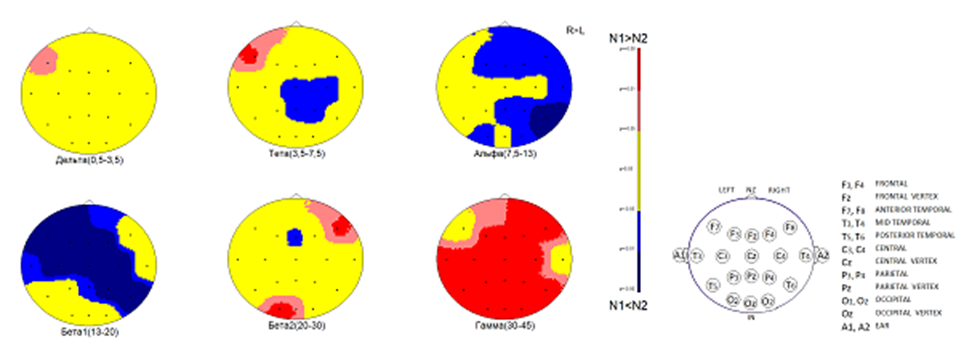
Рис. 2. Испытуемый В.Б. Результаты статистического сравнения спектральной мощности ритмов ЭЭГ. Наблюдаются достоверные изменения в распределении и величине спектральной мощности ритмов во время чтения книги без участия глаз по сравнению с фоновым состоянием. Обозначения: шкала коэффициентов достоверности и схема отведений ЭЭГ в правой части рисунка; ниже каждой карты обозначены ритмы ЭЭГ. Пояснения в тексте
Кроме того, обращает на себя внимание достоверное увеличение спектральной мощности ритмов бета2 (20–30 Гц) и гамма (30–45 Гц) на поверхности коры обоих полушарий в сенсомоторной зоне кожно-мышечного восприятия. При этом увеличение происходит также в правой лобно-височной области и в зрительной коре в затылочных областях обоих полушарий. В настоящее время пока трудно объяснить, какие сигналы о содержании текста книги поступают в правое полушарие, но на эти сигналы, прежде всего, реагируют сенсомоторные зоны в центральных областях коры обоих полушарий и центры зрительного восприятия в сочетании с ассоциированными с ними областями коры.
«Виртуальные глаза» не могут воспринимать и передавать внешние сигналы в зрительные центры коры таким же способом, как и естественные глаза. Но несмотря на это, зрительные образы, воспринимаемые «виртуальными глазами» В.Б., имеют обычный естественный вид, при этом не нарушается даже цветовая гамма. Следует обратить внимание, что во время чтения книги в маске на ЭЭГ одновременно повышается мощность ритмов гамма-диапазона в первичных центрах не только зрительной, но и кожно-мышечной, и слуховой сенсорной модальности (рис. 2). Это даёт возможность предположить возникновение зрительно-слуховых и зрительно-кинестетических синестезий.
Предполагается, что «виртуальные глаза» могут реагировать только на длины волн отражённого объекта, а цветное изображение формируется в зрительных зонах коры.
Ранее было обнаружено, что глаза и точки акупунктуры излучают колебания инфракрасного (ИК) и миллиметрового (КВЧ) диапазонов [20, с. 26-35], а также чувствительны к воздействиям ЭМВ ИК и КВЧ диапазонов [20, с. 26-35; 21]. То, что чтение книги в маске на глазах происходит с участием волн, подтверждает факт снижения эффективности распознавания текста в связи с увеличением длины волн «опорного излучения», которое наблюдается при утомлении [9, 10]. Было показано, что увеличение длины волны опорного излучения более 2 мм является устойчивым признаком утомления испытуемых, владеющих альтернативным видением.
Были также зарегистрированы не известные ранее микроволновые ЭМИ УВЧ/СВЧ-диапазона (от 1,5 до 4,5 ГГц) с мощностью сигналов на уровне -130 дБм –100 дБм (1*е-15 – 1*е-13 Вт), имеющие зональные различия в различных областях головы человека и отсутствующие при измерениях от других участков тела обследуемых. Регистрация микроволновых излучений головного мозга человека открывает новый информационный канал.
Полученные результаты картирования спектральной мощности бета- и гамма-ритмов ЭЭГ дают возможность начать рассмотрение путей поступления информации без участия глаз для построения зрительных образов и восприятия книжного текста. Значительное увеличение мощности этих ритмов в сенсомоторной зоне обоих полушарий во время альтернативного видения по сравнению с фоном можно интерпретировать как усиление кожной чувствительности к восприятию внешних сигналов. Если эти сигналы относятся к восприятию книжного текста или других зрительных объектов, то возможны два пути их дальнейшего преобразования в осознаваемые зрительные образы. Первый возможный путь преобразований – синестезии, который осуществляется за счёт биофизического взаимодействия сенсорных систем разной модальности благодаря особенностям распространения и сенсорного стробирования альфа-ритма [2, с. 19-29; 3, с. 6-12]. Второй возможный путь – ассоциативная память о сочетаниях поступающих внешних сигналов с уже ранее запомнившимися зрительными образами или их деталями.
Программы обучения и тренировки альтернативного видения, разработанные создателями школы В. М. Бронникова, успешно применяются для развития этой способности у слепых и слабо видящих детей, а также у взрослых. Однако для достижения результатов некоторыми учащимися требуется достаточно длительное время – месяцы, иногда годы. В таких случаях для развития необходимых мозговых функций требуется дополнительная информация о состоянии важных для их исполнения нейроструктур и об эффективности взаимодействия между ними.
Существует возможность совершенствовать систему обучения, опираясь на чёткие и стабильные показатели альтернативного (внешнего) видения, которые можно использовать для развития необходимых функций у обучаемых.
Например, на рисунке 3 представлены данные об изменениях спектральных характеристик ЭЭГ во время чтения книги в состоянии альтернативного видения по сравнению с фоном у испытуемого В. Б. (рис. 3 (1)).
Аналогичные исследования, проведённые с участием учащихся (рис. 3 (2, 3, 4)), показывают изменения спектральной мощности ритмов ЭЭГ, сходные с изменениями у В.Б., но отсутствует очень важный признак – нет повышения активности в лобно-височной области левого полушария. Максимальная активность мозга именно в этой области, выраженная в дельта-диапазоне ритмов ЭЭГ (рис. 1 (2)), может порождать излучение инфракрасных ЭМВ, которые, благодаря проницаемости черепа, могут стать источником «опорного излучения» для голографического отображения внешних объектов при альтернативном видении.
С другой стороны, сходство изменений спектральных характеристик ЭЭГ учащихся, несмотря на индивидуальные различия ЭЭГ, говорит о высокой эффективности обучения. Однако возникает задача дальнейшего повышения эффективности обучающих программ для развития определённых функций лобно-височной области левого полушария.
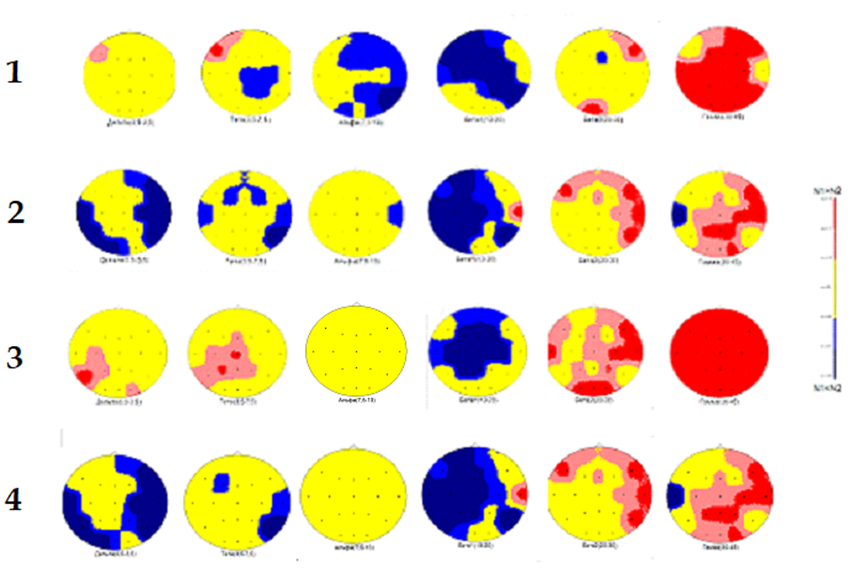
Рис. 3. Сравнение спектральной мощности двух ЭЭГ-записей (критерий Стьюдента) по отдельным диапазонам ритмов (дельта, тета, альфа, бета1, бета2, гамма – указаны под изображением карты). Отведения ЭЭГ как на рис.2. Шкала уровня значимости р со знаком (двухсторонний (N1-N2) расположена в правой части рисунка. Испытуемые: 1) В. Б., 2) О. К., 3) А. Л., 4) Д. Ц. Описание в тексте
Чёткие отличия наблюдаются и в показателях процентного отношения выявленных ЭДИ дельта-ритма при КД ≥ 0.94 к общему количеству ЭДИ. Высший показатель 12% у В.Б., учащиеся имеют более низкие цифры. Это дополнительно показывает большое значение активности лобно-височной области левого полушария для развития альтернативного видения. ЭДИ гамма-ритма, напротив, в процентном отношении имеют разные, в том числе, более высокие цифры, чем у В. Б. Но и при более высоких цифрах плотное распределение ЭДИ наблюдается в других структурах, а в зрительной области затылочной коры количество ЭДИ меньше, чем у В.Б. Это наблюдение ставит следующую задачу повышения эффективности программ обучения: развитие когнитивных способностей, связанных с взаимодействием зрительной системы с ассоциативной памятью и синестезиями.
Альтернативное видение – это сложная реорганизация нейронных сетей, где мозг использует нестандартные сенсорные пути и усиленные когнитивные механизмы. Для достижения этого состояния необходима целенаправленная тренировка интеграции тактильных, проприоцептивных и мнемических сигналов, а также контроль ритмической активности мозга (особенно дельта-, тета- и гамма-диапазонов). Такими способами проявляемая нейропластичность мозга компенсирует отсутствие зрения, перераспределяя ресурсы на тактильные, проприоцептивные и мнемические пути.
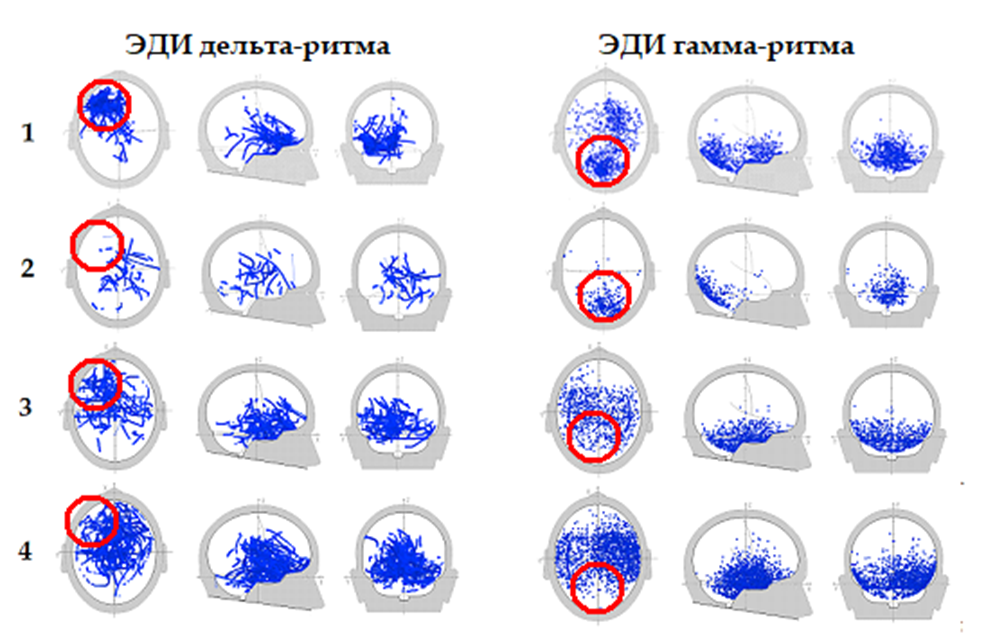
Рис. 4. Распределения ЭДИ дельта и гамма ритмов ЭЭГ в состоянии альтернативного видения. Красной окружностью обозначены области принципиально важной активности для проявления альтернативного видения. Указано процентное отношение выявленных ЭДИ при заданном КД к общему количеству ЭДИ при минимальном КД у испытуемых: 1) В.Б. – дельта 0.5–3.5 Гц, КД ≥ 0.94; 12.0%; гамма 30–45 Гц, КД ≥ 0.96 1.6%; 2) О.К. – дельта 0.5–3.5 Гц, КД ≥ 0.94; 1.5%; гамма 30–45 Гц, КД ≥ 0.96 0.6%; 3) А.Л. – дельта 0.5–3.5 Гц, КД ≥ 0.94; 6.6%; гамма 30–45 Гц, КД ≥ 0.96 2.0%; 4) Д.Ц. – дельта 0.5–3.5 Гц, КД ≥ 0.94 10.6%; гамма 30–45 Гц, КД ≥ 0.96 4.0%
На рисунке 4 представлены распределения ЭДИ дельта- и гамма-ритмов ЭЭГ в поверхностных и глубоких структурах мозга у испытуемого В. Б. и учащихся (рис. 4). Лобно-височная область левого полушария обведена красной окружностью для сопоставления плотности распределения активных центров, обозначенных локализацией ЭДИ. Наблюдается снижение количества ЭДИ в окружности за счёт рассеивания и уменьшения их плотности у обучаемых лиц. Именно центр концентрации ЭДИ дельта-ритма у В. Б. находится в центре окружности. У остальных испытуемых центры концентрации ЭДИ сдвинуты в сторону правого полушария и не соответствуют зонам активности полей Бродмана 11,47,38.
Поэтому испытуемые с распределениями ЭДИ дельта-ритма, как показано на рисунке 4 (2, 3, 4), могут распознавать только отдельные буквы или слова, но не могут читать книжный текст целиком.
Если максимальная концентрация ЭДИ гамма-ритма у В.Б. наблюдается в зрительной зоне коры затылочной области (рис. 4 (1)), то у обучающихся (рис. 4 (2, 3, 4)) концентрация ЭДИ или снижается (рис. 4 (2)), или смещается в другую область коры больших полушарий. Поэтому увеличение количества ЭДИ гамма-ритма у обучаемых (рис. 4 (3, 4)) становится дополнительным «когнитивным каркасом» для альтернативного восприятия.
Нейрофизиологические методы (ЭЭГ) объективно подтверждают существование феномена альтернативного видения у лиц, способных к восприятию внешних объектов и чтению книг без использования глаз, а также у обученных альтернативному видению лиц. Мозг переходит в качественно иное функциональное состояние при его реализации, затрагивающее участие систем восходящей неспецифической активации коры мозга, центры эмоциональной регуляции лимбической системы, системы восприятия различных сенсорных модальностей.
Выводы
1. Альтернативное зрение – способность воспринимать зрительную информацию без использования глаз, подтверждено экспериментальными исследованиями под руководством академика Н. П. Бехтеревой и проф. Ю. П. Пытьева, которые доказали, что это воспроизводимый и изучаемый феномен, а не уникальное явление.
2. В настоящей работе продолжены нейрофизиологические исследования, которые показали, что при альтернативном зрении наблюдаются статистически значимые изменения в ЭЭГ:
- усиление дельта- и тета-ритмов в лобно-височной области левого полушария (зоны Бродмана 11, 38, 47);
- активация гамма-ритмов в зрительной коре и в сенсомоторных зонах обоих полушарий.
Это указывает на перестройку функционального состояния мозга и вовлечение нестандартных нейронных сетей.
3. Левое полушарие (особенно ВА11) играет ключевую роль в организации процесса альтернативного зрения и является дополнительным подтверждением голографической модели восприятия, предложенной проф. Ю. П. Пытьевым. Высокая активность в ВА11 совпадает у В. Б. с областью излучения «опорной волны» (длина волны~1,5–2 мм), которая интерферирует с объектным излучением от книжного текста.
4. При альтернативном зрении активируются первичные центры разных сенсорных модальностей (зрение, слух, тактильность). Это позволяет предположить участие синестезий – преобразования сигналов из одной модальности в другую.
5. Обучение по методу В. М. Бронникова позволяет развивать альтернативное зрение у слепых, слабо видящих и зрячих с использованием оптимального вида голограммы восприятия без участия глаз. Дополнительным объективным показателем голограммы является концентрация активности в левой лобно-височной области (дельта-ритм в сочетании с инфракрасным излучением).
6. Нейрофизиологический мониторинг (ЭЭГ, ЭДИ) позволяет оценивать прогресс в обучении и корректировать программы. Выявленные «эталонные» паттерны активности мозга (у испытуемого В.Б.) могут служить ориентиром для развития способности у других.
7. Остаются нерешённые вопросы. Природа «опорного излучения» (электромагнитное или иное) и механизм его генерации сознанием остаются не до конца изучены. Не ясен точный путь передачи сигналов от «виртуальных глаз» к зрительной коре.
8. Альтернативное зрение – это сложный, но реальный феномен, основанный на нейропластичности, голографических принципах восприятия и сенсорной интеграции. Его изучение открывает новые возможности для реабилитации слепых и понимания скрытых ресурсов человеческого мозга.
