
Соя – важная сельскохозяйственная культура, богатая белками, жирами, микро-и макроэлементами. Спрос на культурные сорта сои неуклонно растет во всем мире. Несмотря на создание урожайных сортов и повышение урожайности, имеют место и потери. Большая часть потерь происходит из-за абиотических стрессовых факторов и составляет около 1/5 валового продукта [3, с.1]. Поэтому, изучение физиолого-биохимических аспектов влияния засухи и засоления характерных для климата Азербайджана, на растение сои является приоритетным вопросом.
Важное значение в селекции устойчивых сортов и форм имеет разработка ранней диагностики устойчивости к засухе и засолению растений на основе физиолого-биохимических показателей стрессоустойчивости.
Материалы и методика
В исследовании использованы образцы 5 сортов культурной сои (Glycine max L.): Браво, Бейсон, Агройол, Регале, Синара, выращенных на опытно- полевом участке Института генетических ресурсов. Степень устойчивости образцов сои определяли по изменению содержания хлорофилла, пролина и активности гваякол-пероксидазы в листьях при засухе и засолении. Для изучения взаимосвязи между устойчивостью генотипов сои к стрессовым факторам и содержанием хлорофилла, в качестве экспериментального материала были взяты образцы 18-20 дневных опытных полевых интактных растений с 6-7 листьями. Растения подвергались стрессу в лабораторных условиях. Для моделирования стресса были использованы растворы сахарозы 8,7% (7 атм) и 1,5% NaCl (11 aтм), соответствующие пределу устойчивости [1, с.61].
Уровень хлорофилла в верхних и средних листьях измеряли с помощью портативного аппарата SPAD-502Plus. Устройство определяло спектральное поглощение в двух диапазонах, и на основании полученных данных рассчитывало индексированное значение содержания хлорофилла в листьях. Определяли процентное соотношение содержания фотосинтетических пигментов опытных вариантов и контроля, и это соотношение было принято за единицу измерения для выявления устойчивых к стрессу образцов: чем выше были показатели, тем образец оценивался как более устойчивый.
Содержание пролина определяли с помощью нингидринового реактива по методу Bates et. аl. [6]. Для этого использовали образцы верхних листьев растений различных вариантов полевого опыта, которые также подвергались воздействию засухи и засоления. Оптическая плотность пролина измерялась на спектрофотометре (UV-3100 PC) при длине волны 520 нм.
Определение активности пероксидазы спектрофотометрическим путем [2, с.42] основан на измерении оптической плотности продуктов, образующихся в ходе реакции окисления гваякола. 200 мг листьев измельчали в небольшом количестве (10 мл) фосфатного буфера (рН 5,4) в фарфоровой чашке в течение 10 минут после измельчения, центрифугировали со скоростью 4000-5000 об/мин. Оптическая плотность реакционной смеси, состоящей из 0,5 мл субстрата (гваякола), 1,5 мл фосфатного буфера, 0,5 мл супернатанта (ферментативного растительного материала), 0,5 мл H2O2, измеряли в течении 1 мин на спектрофотометре (UV-3100 PC) при длине волны 470 нм.
Повторность опытов трехкратная.
Результаты и обсуждения
Содержание хлорофилла (а+b), определяемое методом SPAD, показало, что у контрольных растений этот показатель колебался в пределах 27,2-33,5 (рис.1). Максимум пигментов пришелся на образец сорта Браво, а минимум на экспериментальный вариант Бейсон.
В условиях засухи сумма фотосинтетических пигментов изменялась в пределах 25,7-32,4, максимальный показатель наблюдался в выборке сорта сои Регале, а минимальный у растений экспериментального варианта Бейсон. У опытных вариантов Браво и Бейсон сумма хл (a+b) во время засухи была заметно меньше, соответственно, 5,6 (17%) и 1,5 (5,5%) относительно контроля, а у образца Агройол наблюдалось уменьшение в незначительных количествах (0,6). У опытного варианта Регале было выявлено увеличение показателя хл (a+b) на 4 (14%) по отношению к контролю, а у образца сорта Синара увеличение было незначительным.
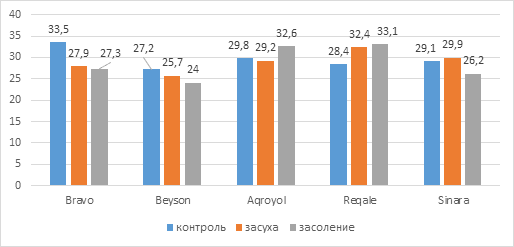
Рис. 1. Содержание хлорофилла (a+b) у образцов сои
Сумма фотосинтетических пигментов под воздействием засоления имела значения в более широком диапазоне 24-33,1. Так, в совокупности у растений сорта Регале наиболее высокий показатель содержания хлорофилла, а у растений образца сои Бейсон самый низкий. Показатели содержания хлорофилла (а+b) в процентном соотношении к контролю составили 116,5% (Регале), 109,4% (Агройол), 90% (Синара), 88,2% (Бейсон), 81,5% (Браво).
Визуальные наблюдения за опытными растениями сортов сои, через сутки после воздействия засухи, соответствующей пределу устойчивости, показало, что растения сортов Браво, Бейсон и, в особенности, Агройол сохранили тургор, листья не пожелтели и не увядали. В то время как в опытном варианте Регале наблюдалась небольшая потеря тургора, эффект стресса более четко наблюдался у сорта Синара. Физиологические наблюдения показали, что растения сорта Синара не выдержали 48-часового воздействия засухи. В отличие от стресса засухи, более мягкое воздействие засоления на экспериментальные образцы привлекло внимание. Так, что даже 48-часовое воздействие не привело к гибели растений ни одного из вариантов.
Опираясь на полученные результаты, можно отметить, что положительной корреляции между содержанием хлорофилла и устойчивостью растений не наблюдается. В то время как воздействие стрессовых факторов засухи и засоления вызывают деградацию фотосинтетических пигментов в опытных вариантах сортов Браво и Бейсона, эти образцы сортов сои продемонстрировали устойчивость к 24-часовому воздействию стресса засухи. В то время, как вариант сои Синара, несмотря на увеличение содержания хлорофилла после 1-дневного воздействия засухи, был физиологически ослаблен, в листьях наблюдалась частичная потеря тургора и увядание, а 48-часовое воздействие стресса приводило к его гибели.
В качестве общей физиологической ответной реакции у растений, подвергшихся воздействию различных абиотических стрессовых факторов, происходит повышение содержания пролина. Определение содержания свободного пролина и активности гваякол-пероксидазы в листьях опытных вариантов сои после 24-часового воздействия солевого стресса показало, что активность фермента снижалась, в то время как содержание аминокислоты пролин повышалось во всех вариантах (рис.2). После суток воздействия солевого стресса содержание пролина колебалось в пределах 0,151 -0,318 мкМ/мг, причем, эти показатели составили 342% (Синара), 220% (Браво), 487% (Бейсон и Агройол), 291% (Регале) по отношению к контролю. Увеличение содержания пролина может быть следствием его синтеза de novo, снижением деградации, утилизацией в меньшей степени и деградацией белков. Определение активности фермента после 24-часового воздействия солевого стресса на образцы сои показало, что за исключением варианта Синара, у остальных вариантов опыта активность гваякол-пероксидазы снизилась. Активность фермента в растениях, помещенных на 1 сутки в соленую среду, изменялась в интервале 0,095 – 0,284 мкМ/мг и составила 43% (Агройол) и 145% (Синара), относительно контроля.
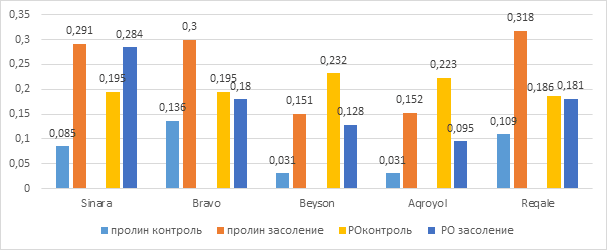
Рис. 2. Содержание пролина в листьях растений сои и активность гваякол-пероксидазы при 24-часовом воздействии солевого раствора (мкМ/мг)
Для поддержания жизнеспособности в неблагоприятных условиях у растений создан двухкомпонентный эффективный антиоксидантный механизм: ферментативные компоненты, состоящие из семейства SOD, каталазы и пероксидазы, и неферментативные компоненты, состоящие из аскорбиновой кислоты, α-токоферола, каротиноидов, флавоноидов, пролин-осмолита. Эти два компонента вместе выполняют такую важную функцию, как нейтрализация АФК (активных форм кислорода), которые образуются в значительных количествах во время стресса и могут представлять серьезные угрозы для нормального выживания растительного организма [7]. В нашем исследовании, для всех опытных образцов, за исключением сорта Синара, можно судить о взаимодействии компонентов системы антиоксидантной защиты, основываясь на взаимной изменчивости их количества и активности. Так, у образца сорта Браво влияние стресса засухи привело к увеличению cодержания свободного пролина в 2,2 раза, а активность гваякол-пероксидазы-к снижению в 1,08 раз. У образца сорта Бейсон содержание пролина увеличилось в 4,8 раза, активность фермента снизилась в 1,81 раз, в вариантах Агройол и Регале, соответственно, содержание пролина увеличилось в 4,8 и в 2,9 раза, а активность фермента снизилась в 2,3 раза у сорта Агройол и в незначительных количествах у сорта Регале.
В последнее время все больше внимания уделяется антиоксидантной способности пролина. Особенности его строения позволяют рассмотреть возможность непосредственной инактивации радикальных форм кислорода. Как известно, пролин играет большую роль в инактивации гидроксильного радикала, а также участвует в детоксикации перекиси водорода, когда содержание пролина в клетке в больших количествах [10, с.37; 11, с.9]. Поскольку ферментные антиоксиданты могут нейтрализовать перекись водорода, супероксидные радикалы и синглетный кислород, такой механизм является удобным способом защиты от свободных радикалов, так как они не могут нейтрализовать гидроксильные радикалы, а количество пролина в компартментах клетки достаточно велико. С другой стороны, локализация значительного содержания свободного пролина в цитоплазме также свидетельствует о том, что он защищает антиоксидантные ферменты от воздействия, которое может привести к различным повреждениям и денатурации [4, с.61-62; 5, с. 42; 9, с.1587].
Принимая во внимание тот факт, что пролин многофункционален и выступает в качестве осморегулятора в формировании адаптации растений к засухе и засолению, его накопление в больших количествах в клетке приводит к повышению внутриклеточной осмолярности и к поддержанию водного баланса, водного потенциала и, что важно, тургора в условиях дефицита воды [8, с.18]. Таким образом, проведенное нами исследование показало, что восстановление нарушенного водоснабжения в тканях растений при раннем стрессе создало условия для сохранения фотосинтетических пигментов, хотя и в незначительных количествах деградирующих. Несмотря на то, что по количеству фотосинтетических пигментов в стрессовых условиях образец сои сорта Регале доминирует, прямой корреляции между количеством хлорофилла и пролина не наблюдалось. Согласно литературным данным, для увеличения содержания пролина у устойчивых сортов требуется более сильное и (или) длительное воздействие стресса [5, с.44]. Учитывая результаты визуальных наблюдений и анализов по определению содержания пролина можно отметить сорта Бейсон и Агройол более устойчивыми.
Предположительно, компоненты системы антиоксидантной защиты взаимодействуют в нейтрализации радикальных форм кислорода.
