
Введение
В связи с повышением общественного интереса к творческой деятельности человека возросло количество научных исследований, посвящённых расширению сферы восприятия, в частности, развитию синестезий.
В переводе с греческого языка синестезия – это одновременное ощущение, совместное чувство. Иными словами, это нейрофизиологический феномен, при котором раздражение в одной сенсорной системе ведёт к автоматическому, непроизвольному отклику в другой сенсорной системе.
Кроме того, синестезии в значительной степени расширяют и обогащают восприятие человека за счёт комплексного настроя органов чувств на фоне соответствующих эмоциональных переживаний. В связи с этим, значительно возрос интерес к синестезиям не только у психологов, но и у специалистов, занимающихся развитием творческих возможностей человека в области культурологии, музыковедения, живописи и других различных видов искусства.
Современные подходы к изучению синестезий значительно расширены. К настоящему времени выделены и описаны отдельные типы синестезий [1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11].
Наиболее устойчивые синестезии в виде разномодальных ощущений в ответ на моносенсорное раздражение является редким явлением (в 1….4 % случаев). Оно имеет физиологическую основу, которая характеризуется непроизвольностью протекания, врожденностью и генетической наследственностью [1, 3, 8, 11].
Семантические синестезии могут возникать посредством извлечения смысла при действии синестетического стимула. В таком случае синестезия становится принципиально семантическим феноменом и понимается как некий механизм взаимоперевода информации из одной модальности в другую. Данный вид синестезии связан с семантическими структурами, универсален и проявляет эмоционально-оценочный характер [2, 3, 7, 9, 10].
Наконец, синестезия может возникнуть, как естественная сознательная способность человека соотносить объекты различных модальностей, выражать межчувственную ассоциацию, выработанную в культуре и приобретенную в процессе социализации [3, 5, 7].
Синестезии могут быть более разнообразными. Шон Дей [11] проанализировал 175 случаев смешения чувств и насчитал девятнадцать вариантов. Выявив множество неверных трактовок синестезий, ошибок и заблуждений, автор выдвинул обоснованные положения, касающиеся четырех различных подгрупп проявлений синестезии: врожденной синестезии, посттравматической, интоксикационной и синестезии измененного состояния сознания. При этом он отметил, что наибольшую часть (69%) составляют случаи цветозвукового восприятия.
Однако отдельные авторы [6], рассмотрев немногие из смыслов, употребляемых в термине «синестезия», пришли к выводу, что не стоит пока искать какого-то единого определения, чтобы не потерять самое интересное – те интенсивные дискуссии и богатые контексты, которые сопровождают такой не до конца еще определенный концепт как синестезия.
В настоящем исследовании синестезии рассматриваются как способность преобразования сигналов, поступающих в мозг по сенсорным каналам одной модальности, в образы другой сенсорной модальности. Подобные синестезии могут вносить качественные изменения в систему восприятия, поскольку могут переводить неосознаваемые ощущения одних органов чувств в осознаваемые другой, более развитой у конкретного человека системой восприятия [4, 12]. Данный тип синестезии характерен для экстрасенсорного восприятия и может возникать на фоне особых эмоциональных состояний, например, таких как эмпатия (сопереживание).
Прежде чем приступить к исследованию синестезий, необходимо было вначале выявить и представить алгоритм считывания сигналов активности мозга в зонах первичного восприятия отдельных видов чувствительности (зрительной, слуховой, соматосенсорной). Это давало бы возможность определить явления, связанные с взаимодействием этих зон, сведения о которых позволяли описать собственно синестезии. Предполагалось, что критерием оценки поступления специфических сигналов в первичные зоны и показателем межзональных взаимодействий, могут служить параметры спектральных характеристик альфа-ритма ЭЭГ.
На особенности частотного спектра альфа-ритма ЭЭГ первым обратил внимание Н.Винер [13]. Он особо отметил то, что преобладающая часть мощности спектра ритмов мозга сосредоточена в окрестности частоты 10 Гц. Определив в энцефалограмме конкретного человека крутое падение мощности на частоте 9,05 Гц, достаточно устойчивое во времени, он отметил, что резкая линия изменения частоты эквивалентна точным часам, обеспечивающим механизмы стробирования. При этом стробирование можно рассматривать, как один из механизмов в развитии системы саморегуляции. Это позволяет осуществлять комбинирование действий в интервалах времени, определяемых стробированием.
Таким образом, описанное свойство альфа-ритма способствует проявлению его частотной специфичности для каждой отдельной зоны сенсорной модальности и возможности взаимодействия между разномодальными зонами первичного восприятия с использованием эффекта стробирования.
В исследовании необходимо было также учитывать ведущий тип восприятия у каждого человека. Авторы работы исходили из того, что:
- визуалы – это люди, у которых доминирующим каналом восприятия является зрение;
- аудиалы – люди, у которых доминирующей является слуховая система обработки информации;
- кинестетики – люди, воспринимающие мир на ощупь, доминирующей у них является чувственная информация прикосновения, вкус, запах, ощущение текстур, температуры;
- дискреты – лица, восприятие которых, прежде всего, связано с логическим построением внутреннего диалога.
У таких лиц доминирующая система восприятия является, как правило, пусковой, ведущей, как для ментальных процессов, так, возможно, и для синестезий.
Однако у большинства людей доминирование того или иного типа восприятия является неустойчивым и в значительной степени зависит от текущей ситуации и психологического настроя.
Таким образом, для выполнения перечисленных задач, направленных на изучение синестезий, необходимо ответить на следующие вопросы, имеющие непосредственное отношение к структуре исследования.
- Какая частота альфа-ритма характерна для каждой зоны первичного восприятия?
- Где располагаются источники происхождения альфа-ритма каждой корковой зоны первичного восприятия?
- Как глубинные источники происхождения альфа-ритма проецируют активность в кору головного мозга?
- Какова роль альфа-ритма в восприятии сигналов и в формировании образов разных ощущений?
- Каковы особенности спектральных характеристик альфа-ритма и их распределения на поверхности коры у визуалов, акустиков, кинестетиков, дискретов? Как у них локализуются источники происхождения альфа-ритма?
- Является ли постоянно доминирующая в коре мозга зона первичного восприятия (аудиала, визуала, кинестетика, дискрета) проявлением синестезий по отношению к остальным зонам восприятия ощущений?
- Какое влияние оказывают эмоции, возбуждаемые музыкальными мелодиями, на возникновение синестезий, оцениваемых по спектральным показателям альфа-ритма?
- Каковы основные объективные характеристики синестезий по данным спектральных характеристик ЭЭГ и зонам локализации эквивалентных дипольных источников происхождения альфа-активности?
Методика
В исследовании участвовала группа испытуемых, занимающихся интеллектуальным трудом (25 чел., из них 15 женщин и 10 мужчин), в возрасте от 30 до 60 лет.
Для получения ответов на выше поставленные вопросы 1….6 было достаточно провести у испытуемых стандартную фоновую регистрацию ЭЭГ в состоянии психической и мышечной релаксации с закрытыми глазами.
Это предварительное исследование позволило создать методический инструмент в виде комплекса объективных показателей альфа-ритма для изучения синестезий.
Вопрос о возможности возникновения синестезий, если они не являются врождёнными, решался с помощью тестирования эмоциями при прослушивании любимых музыкальных мелодий.
Для тестирования и оценки эмоционального состояния была использована система акустического воздействия. Предполагалось, что непосредственное эмоциональное воздействие на испытуемых происходит на уровне слухового восприятия информации намного точнее, нежели с опорой, например, на мимические или иные изображения эмоциональных значений [14]. Поэтому предпочли использовать музыкальные мелодии с определённым эмоциональным содержанием, чтобы запустить у испытуемого переход в соответствующее эмоциональное состояние. Этот процесс более вероятен при прослушивании любимых мелодий. Возможно, уровень перехода к эмоциональному сопереживанию мелодии будет зависеть от исходного развития этого чувства или иначе – эмпатии. Речь идёт о расширении сферы восприятия.
В качестве наиболее адекватных методов получения объективных данных о состоянии первичных корковых зон восприятия ощущений и о динамике эмоциональных состояний сознания были выбраны нейрофизиологические способы регистрации электроэнцефалограммы (ЭЭГ).
Регистрацию ЭЭГ проводили на 24-канальном нейровизоре NVX24 производства ООО "Медицинские Компьютерные Системы" с использованием стандартных монополярных отведений в соответствии с принятой международной схемой 10-20 [15].
Исследования проводились в стандартных условиях, одинаковых для испытуемых. Биопотенциалы мозга регистрировали при закрытых глазах в состоянии бодрствования с психической и мышечной релаксацией и в состоянии эмоционального возбуждения при прослушивании любимой мелодии или музыки, содержащей одну из базовых эмоций. Для исключения влияния шумовых помех или случайных звуковых сигналов были использованы наушники.
Для оценки изменений ЭЭГ при тестовых испытаниях использовали процедуру сравнения с ЭЭГ в состоянии фона, то есть психической и мышечной релаксации с закрытыми глазами (для снижения уровня сигналов, поступающих от тела и через зрительный канал). В этом случае фоновые показатели активности мозга становились началом отсчёта, так как условия регистрации (освещение, уровень шумовых помех, температура воздуха, состояние организма и т.д.) оставались одинаковыми как в фоне, так и при выполнении тестов. Поэтому отличия в показателях, которые возникали при выполнении тестов, были отнесены непосредственно к тестируемым процессам. Поскольку функциональная изменчивость ЭЭГ характерна и в пределах нормы, то во внимание принимались только статистически достоверные отличия.
Рис. 1. Стандартное расположение электродов на поверхности головы по схеме 10/20
Система обработки данных
Основная цель обработки данных регистрации ЭЭГ заключалась в топической оценке спектральных характеристик альфа-ритма и нейрокартировании с частотным шагом 1 Гц в пределах всего диапазона альфа колебаний 7 Гц…13 Гц. Рассматривалась также локализация центров электрической активности, ответственных за происхождение отдельных частот альфа-ритма. Для этого использовали пакеты компьютерных программ, обеспечивающих спектральный анализ регистрируемых сигналов и топографическое картирование спектральных характеристик ЭЭГ [16]. В результате получали распределение показателей мощности частотного спектра по отдельным областям на поверхности полушарий мозга. Использован статистический анализ данных для определения достоверности полученных как индивидуальных, так и групповых изменений.
С целью выявления наиболее выраженных реакций при выполнении индивидуальных проб вычисляли локализацию и распределение эквивалентных дипольных источников (ЭДИ) происхождения электрической активности в глубинных структурах мозга. Для этого определяли и отслеживали в объёме мозга человека локализацию ЭДИ, вычисляемых в каждый момент времени, равный дискретному интервалу отсчёта данных при вводе в компьютер. Программа использовалась для того, чтобы проследить возникновение центров электрической активности в структурах мозга при эмоциональных состояниях. Вопрос о соответствии локализации источников в глубинных структурах мозга той биоэлектрической активности, которая регистрировалась на поверхности головы, решался специальным алгоритмом, в котором рассматривались электрические свойства мозга как объёмного проводника [17, 18].
Результаты и обсуждение.
Распределение спектральной мощности альфа-ритма по зонам коры позволяет определить доминирующий тип восприятия и синестезии.
Но большинство людей имеют неустойчивые доминанты, зависимые от оценки ситуации в условиях меняющейся среды и психологического настроя. Поэтому у испытуемых контрольной группы в состоянии психической и мышечной релаксации наблюдается устойчивое равновесие активности первичных зон сенсорного восприятия. Оно выражается в соответствии между частотой альфа-ритма, максимальной мощностью и расположением в соответствующей зоне поступления первичных сенсорных сигналов.
В норме амплитуда альфа-ритма отражает состояние готовности к повышению функциональной активности данной области коры мозга. Модуляция альфа-ритма связана с активностью ядер таламуса и выраженностью таламо-кортикальных связей [22].
Принимаем во внимание распределение максимумов спектральной мощности альфа-ритма у испытуемого в состоянии спокойного бодрствования при закрытых глазах. Определяем частоту альфа-ритма, характерную для каждого центрального анализатора органов чувств.
Известно, что альфа-ритм с максимальной мощностью над сенсомоторной корой (роландический или мю-ритм) имеет более низкую частоту 7-9 Гц, чем альфа-ритм с максимальной мощностью в затылочной (зрительной) области коры 9-13 Гц. Область сенсомоторной коры включает первичный сенсорный центр кинестетического восприятия, а в затылочной области находится первичный сенсорный центр зрительного восприятия.
В височных зонах первичного восприятия звуковых сигналов у отдельных индивидуумов в состоянии покоя увеличивалась мощность не только на частоте 12-13 Гц, но и на частоте 7-8 Гц. Возможно, это связано с наличием двух потоков акустических сигналов и раздельных центров восприятия. Первый – сформированный благодаря воздушной проводимости звука через наружный слуховой проход, барабанную перепонку и цепь слуховых косточек; второй – благодаря тканевой проводимости через ткани черепа.
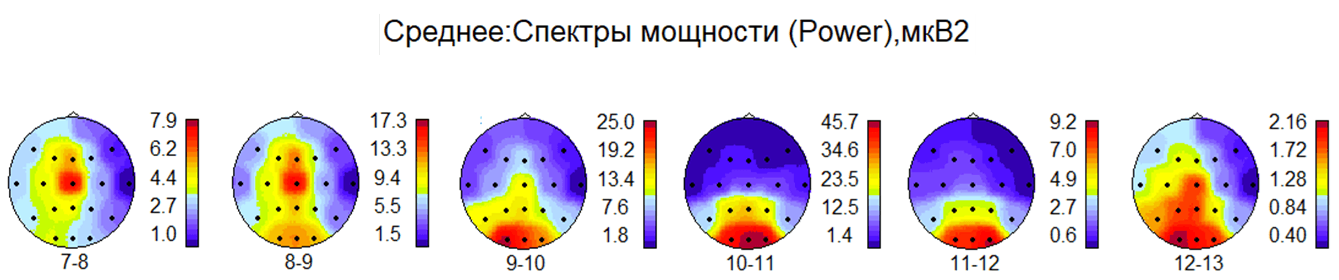
Рис. 2. На рисунке представлены данные группового анализа спектральной мощности альфа-ритма в контрольной группе. Усреднённые максимумы спектральной мощности альфа-диапазона распределились по зонам сенсорного представительства в соответствии с их функциональным значением, т.е. максимум мощности ритма 7...9 Гц находился в сенсомоторных зонах, а ритма 9...12 Гц – в зрительных зонах коры. Первичные зоны слухового восприятия в височных областях имели повышенную мощность альфа-ритма на частоте 12-13 Гц. (Карты коры мозга ориентированы: лоб – сверху, затылок – снизу, справа – шкалы величин мкВ2, внизу – частота альфа-ритма в Гц)
Для подтверждения принадлежности альфа-ритма определённой частоты к передаче сигналов соответствующих видов ощущений была проверена локализация эквивалентных дипольных источников (ЭДИ) альфа-колебаний.
На рис. 3 представлены результаты анализа на примере полученных данных испытуемого И.(контрольная группа). Проведено сопоставление локализации ЭДИ альфа-ритма в глубинных структурах мозга с распределением спектральной мощности альфа-ритма на поверхности коры. Рисунок показывает, что альфа-ритм частотой 7-8 Гц имеет максимальную мощность в центральной области коры мозга, где представлены первичные зоны сенсомоторного восприятия. ЭДИ альфа-ритма данной частоты локализуются преимущественно в полях по Бродману, имеющих отношение к сенсомоторному восприятию и регулированию движений, запоминанию и связям с центрами регуляции эмоций. Кроме того, наблюдается высокая плотность ЭДИ в ядрах таламуса – основных структурах переключения потоков сенсомоторных сигналов. Важно обратить внимание на направленность векторов ЭДИ: в глубинных структурах мозга эти вектора направлены в сторону центральных областей коры с сенсомоторными зонами первичного восприятия и этим обуславливают повышение спектральной мощности именно в этих зонах.
Распределение мощности альфа-ритма частотой 8-9 Гц в основном повторяет картину максимума в центральных областях мозга. Однако увеличивается мощность ритма и в затылочных областях. ЭДИ этого ритма распределяются в аналогичных полях Бродмана. Кроме того, возникают отдельные ЭДИ в зрительной коре затылочной области. А также ЭДИ этого ритма распределяются в структурах системы восходящей неспецифической активации мозга. Вектора ЭДИ в глубинных структурах направлены одновременно к зонам центральной области коры мозга и в меньшей степени к зонам затылочной.
Рассмотрение спектральных характеристик альфа-ритма частотой 9-10-11-12 Гц позволяет получить убедительные сведения в пользу преимущественного распределения максимумов в затылочной области коры. ЭДИ этого ритма, особенно на частоте 9-10-11 Гц, локализуются в повышенном количестве в зонах первичного зрительного восприятия (в полях по Бродману 17,18,19). Они также широко представлены не только в ядрах таламуса, переключающих сенсомоторные сигналы, но и в лимбической системе, в структурах основания мозга, которые отвечают за эмоциональный контроль и запоминание. Вектора ЭДИ, в основном, направлены к поверхности зрительной коры.
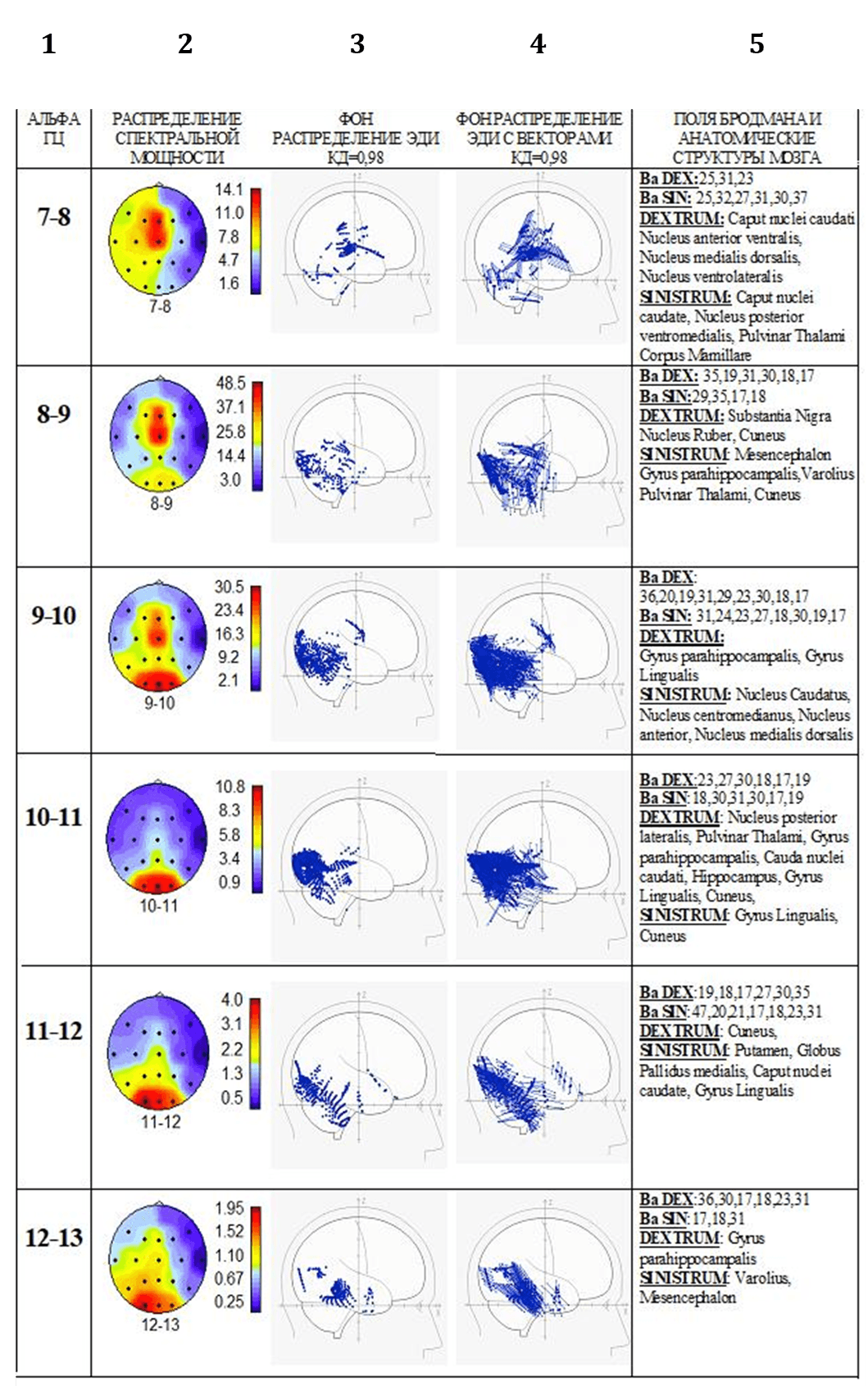
Рис. 3. Испытуемый И. Обозначения столбцов: частота анализируемого альфа ритма (1), пример сопоставления данных распределения спектральной мощности альфа ритма на поверхности мозга (2), локализации ЭДИ происхождения альфа волн (3), локализация тех же ЭДИ с векторами (4), анатомическое обозначение локализации ЭДИ (5). Подробное описание в тексте
Несколько иначе представлена активность альфа-ритма частотой 12-13 Гц. Распределение спектральной мощности этого ритма показывает не только максимум в зрительной коре, но и в височной области, где расположены зоны первичного слухового восприятия. Концентрация ЭДИ этого ритма приближена к внутренним структурам височной области. Глубинные ЭДИ расположены в стволовых ядрах системы восходящей неспецифической активации коры и в лимбической системе, контролирующей эмоции и свойства памяти. Вектора ЭДИ направлены к поверхности как височных, так и затылочных зон коры мозга. В итоге наблюдаются выраженные соотношения между частотой альфа ритма и модальностью сигналов. Альфа ритм частотой 7-9 Гц относится к сенсомоторному, 9-12 Гц – к зрительному, 12-13 Гц – к слуховому восприятию.
Таким образом, были не только подтверждены известные данные о зонах первичного восприятия коры мозга, но и определены конкретные функциональные параметры рассматриваемой нейросистемы в виде частотного стробирования в диапазонах альфа-ритма, которые могут быть использованы как алгоритмический инструмент для изучения более сложных нейрофизиологических процессов. Предполагается, что этот инструмент может быть применим для получения объективных данных о свойствах сознания человека и о некоторых психофизиологических функциях. В данной работе рассматривается конкретное применение для изучения синестезий.
На рис.4. представлены усреднённые по группам распределения спектральной мощности альфа-ритма в коре мозга. Эти распределения в каждой группе имеют свои особенности, но общим является то, что у лиц с доминирующим типом восприятия (зрительным, слуховым или кинестетическим) альфа-ритм распределяется и имеет максимум мощности в доминирующей зоне коры на всех частотах при всех видах сигналов.
В группе аудиалов (Рис.4 АУДИАЛ) максимумы мощности альфа ритма наблюдаются в височных, теменных и затылочных областях в обоих полушариях. В эту группу вошли лица, ежедневно творчески занимающиеся музыкой, прослушиванием и созданием новых мелодий.
Можно предположить, что постоянная творческая деятельность стимулирует именно те системы мозга, которые обеспечивают наиболее широкое восприятие специфических сигналов творчества, восходящие системы неспецифической и эмоциональной активации коры мозга, многочисленные связи и взаимодействия активных центров, в состоянии готовности которых наиболее выражена мощность альфа-ритма.
Особенностью данной группы является повышенная мощность альфа-ритма на всех рассматриваемых частотах во всех первичных центрах сенсорного восприятия. При этом могут ярко проявляться аудио-визуальные синестезии.
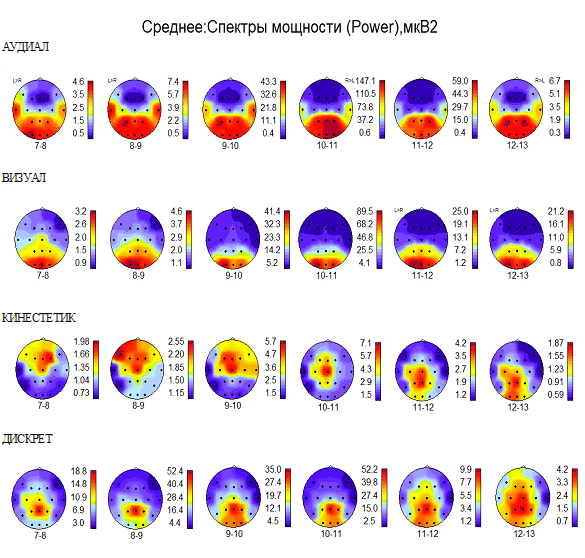
Рис. 4. Усреднённые по отдельным группам аудиалов, визуалов, кинестетиков, дискретов распределения спектральной мощности альфа-ритма. Подробности в тексте
Очевидно также, что альфа-ритм частотой 9-11 Гц имеет отношение к зрительному восприятию и у большинства испытуемых показывает максимальную мощность в зрительной зоне затылочной коры. Однако если одновременно альфа-ритм более низкой частоты 7-9 Гц и более высокой 11-13 Гц также имеет максимум мощности в затылочной области коры, то это означает возможность возникновения кинестетико-визуальных или аудио-визуальных синестезий (Рис.4 ВИЗУАЛ). Такие синестезии могут возникать у визуала, в которых зрительные образы могут сопровождаться более выраженными телесными или слуховыми ощущениями. В этом случае альфа-ритм частотой 7-9 Гц способствует передаче и распознаванию кинестетической, а частотой 12-13 Гц – слуховой информации.
Так, если альфа-ритм частотой не только 7-9 Гц, но и 9-11 Гц, а также 11-13 Гц показывает максимальную мощность в центральных сенсомоторных областях коры мозга, то следует говорить о синестезиях кинестетика (Рис.4 КИНЕСТЕТИК). Это означает, что телесные ощущения кинестетика могут сопровождаться зрительными образами или слуховыми сигналами, потому что альфа-ритм частотой 9-11Гц формирует канал передачи зрительной информации, а альфа-ритм частотой 12-13 Гц – слуховой информации. При этом зрительные и слуховые образы могут быть более яркими и сильными, чем телесные ощущения.
Дискреты – лица, восприятие которых, прежде всего, связано с логическим построением внутреннего диалога (Рис.4 ДИСКРЕТ) имеют повышенную мощность альфа-ритма всех рассматриваемых частот в ассоциативных теменных зонах коры обоих полушарий.
Возникает вопрос, каким образом формируются каналы передачи сенсорной информации, способствующие возникновению синестезий и усиливающие зрительные, слуховые, кинестетические ощущения?
Можно допустить, что альфа-ритм создаёт эквивалентные токовые диполи, вектора которых направлены в слуховую, зрительную, ассоциативную или теменную область коры. С помощью диполей сканирует их и преобразует слабо выраженные или даже неосознаваемые сигналы в яркие и чёткие образы той модальности, с которой ассоциируется сам альфа-ритм.
Подобные рассуждения можно применить по отношению к визуалам, аудиалам, кинестетикам, дискретам. Для того чтобы проверить данное предположение, следует уточнить локализацию ЭДИ в центрах первичного сенсорного восприятия и направленность их векторов в какой-либо другой центр восприятия для преобразования сенсорных сигналов. Решение такой задачи показано на примере нейрофизиологических исследований с представителями каждого из доминирующих типов восприятия.
На рис.5. представлены карты поверхности мозга с распределениями спектральной мощности альфа-ритма с шагом 1 Гц (рис.5.А.1.2.3.4.5.6.) и локализацией эквивалентных дипольных источников его происхождения (рис.5.В.1.2.3.4.5.6.).
У аудиала (рис.5.I.АУДИАЛ ФОН) мощность альфа-ритма на всех частотах увеличена в височных областях в первичных зонах слухового восприятия, а также в ассоциативных теменных областях и в первичных зонах зрительного восприятия в затылочной области. ЭДИ с высоким уровнем дипольности (КД=>0,98) локализуются в височных долях, поясной извилине, вторичных и третичных, а также в ассоциативных зонах сенсорного восприятия коры больших полушарий и в глубинных структурах – гиппокампе, хвостатом ядре, таламусе, среднем мозге. ЭДИ альфа-ритма 7-8 Гц присутствуют в первичной зоне соматосенсорного восприятия и в ассоциативных зонах коры – в полях Бродмана 2,5,7, однако вектора эквивалентных токовых диполей этого альфа ритма направлены в сторону теменной, затылочной и височной областей. Такая направленность векторов ЭДИ создаёт дополнительные возможности сканирования и переноса сигналов эфаптическим путём из одной первичной зоны восприятия в первичный центр другой чувствительности [19]. В данном случае усиливаются возможности создания сложных образов, включающие сигналы соматосенсорной, слуховой и зрительной систем восприятия.
По-видимому, для возникновения синестезии – преобразования сигналов одной модальности в чувственный образ другой модальности, например, слухового образа на основе только зрительных или только соматосенсорных сигналов необходимы дополнительные условия.
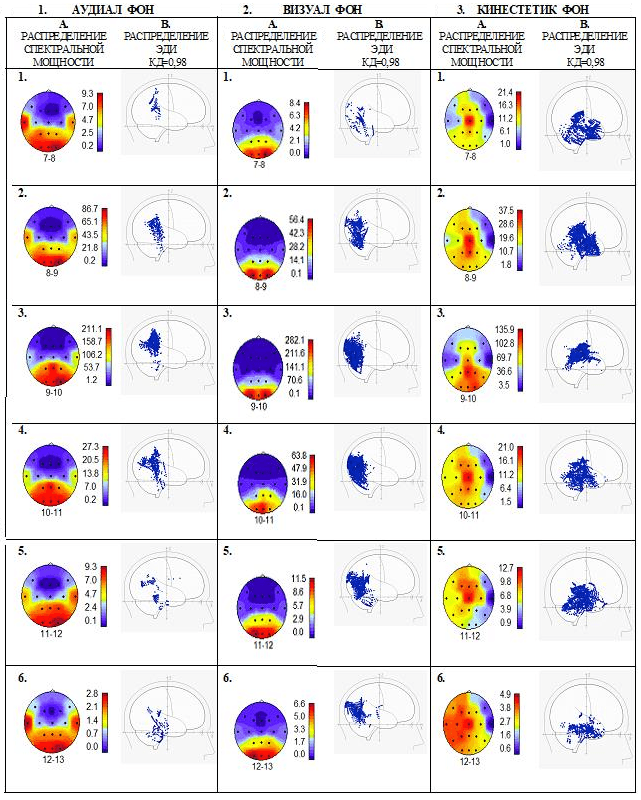
Рис. 5. Примеры анализа ЭЭГ отдельных испытуемых с доминирующим типом восприятия в состоянии спокойного бодрствования, психической и мышечной релаксации (I.,II.,III.). Представлены спектральные характеристики (А.) и локализации ЭДИ (В.) альфа-ритма с разницей частоты шагом в 1 Гц (1,2,3,4,5,6). Обозначения на картах коры мозга ка на рис.2. Подробности см. в тексте
Альфа-ритм визуала (рис.5.2.ВИЗУАЛ ФОН) имеет максимальную мощность в зрительных зонах затылочной коры на всех частотах от 7 Гц до 13 Гц. Необходимо отметить, что альфа-ритм частотой 9-10 или 10-11Гц у всех наблюдаемых испытуемых, независимо от ведущего типа восприятия, имеет максимальную мощность, которая локализуется в доминирующем сенсорном центре. Это соответствует представлению о том, что в организме человека самыми мощными каналами передачи информации являются каналы зрительной системы, и частота альфа-ритма от 9 Гц до 11 Гц связана с частотой зрительного восприятия. Возникает вопрос: почему альфа-ритм частотой 7-8 Гц, характерной для альфа-ритма сенсомоторной зоны, у визуалов имеет максимум мощности в зрительной зоне? Как это может отражаться на телесных ощущениях визуалов? Предполагается, что ответ можно найти в системах взаимодействия альфа-ритма между центральными анализаторами органов чувств в коре головного мозга и в подкорковых структурах.
У визуала основное количество ЭДИ (КД=>0,98) распределено в первичной и вторичной зоне зрительного восприятия. Особенно плотная локализация в этих зонах наблюдается на частоте альфа-ритма 9-10 Гц и 10-11 Гц. Но максимальное скопление ЭДИ альфа-ритма частотой 7-8 Гц наблюдается в поясной извилине. При этом проявляется однонаправленность векторов ЭДИ в сторону зрительной коры мозга. Получается, что альфа-колебания на частоте сенсомоторной зоны могут сканировать зрительную кору и преобразовывать зрительную информацию в телесную. В определённых условиях, когда передача по нейронным путям сигналов от органа чувств будет иметь подпороговое значение и не осознаваться, можно допустить, что анализатор более развитой модальности ощущений с помощью эфаптической связи отсканирует информацию и создаст осознаваемый образ. В этом случае можно говорить о синестезии, то есть о преобразовании сигналов восприятия одной модальности ощущений в образы другой модальности.
У кинестетика (рис.5.3.КИНЕСТЕТИК ФОН) максимум мощности альфа-ритма наблюдается на всех частотах в сенсомоторной зоне в центре коры мозга, несколько смещаясь в ассоциативную зону теменной области только на частоте 9-10 Гц.
ЭДИ с высоким уровнем дипольности (КД=>0,98), включая КД=>0,99, имеют широкое распространение. Они распределены во вторичных, третичных и ассоциативных зонах восприятия различных видов чувствительности в коре больших полушарий и в поясной извилине, а также в структурах лимбической системы, регулирующих эмоциональное состояние и процессы запоминания, в системах восходящей неспецифической активации мозга и эндокринной регуляции. Большинство ЭДИ локализуются компактно в центральных структурах мозга на некотором удалении от зрительной зоны коры. Но вектора большей части ЭДИ направлены в сторону зрительных зон затылочной области и сенсомоторной зоны центральной области. Известно, что в создании сложных образов на основе зрительной и соматосенсорной информации большое значение имеет поле Бродмана 7. Оно является третичным проекционным полем, ассоциативной зоной в месте перекрытия различных анализаторов. Здесь происходит интеграция различных сигналов, формирование соответствующих ощущений, осуществление сложных аналитико-синтетических функций. Колебательные процессы альфа-частотой 9-12 Гц. могут преобразовывать даже слабые неосознаваемые сигналы сенсомоторной зоны в яркие зрительные образы, определяемые как синестезии.
Необходимо отметить, что зарегистрированные с поверхности головы колебания электрических потенциалов интегрально отражают активность мозга, а результаты вычислений эквивалентных дипольных источников (ЭДИ) на основе ЭЭГ отражают эту активность на следующем шаге интеграции деятельности мозга в целом. Поскольку концентрация ЭДИ связана не только с активностью близ расположенных нейронов, а является результатом целостной деятельности мозга, управляемой сознанием, то можно предполагать, что наблюдаемые явления концентрации ЭДИ в большей степени связаны именно с сознанием. Распределение ЭДИ в пространстве межполушарной области отражает степень симметричности участия полушарий мозга в деятельности, связанной с пространственно-временным структурированием активной среды.
Отметим также, что в глубинных структурах мозга и в коре локализуются ЭДИ с более низким уровнем дипольности, чем те, которые конкретно рассматриваются в данной работе. Эти многочисленные ЭДИ, в целом, могут влиять на результаты исследований как общий шумовой фон, но они поддаются начальному запуску тех ЭДИ, которые имеют максимальный уровень дипольности КД=>0,98. При таком условии ЭДИ из разряда шума не только рассеивают, но и подкрепляют динамику поведения активности соответственно высокому уровню дипольности.
Таким образом, проведённая работа позволяет получить некоторые представления о способах обнаружения и о возможности развития синестезий. Осталось ответить на вопрос о том, какое влияние оказывают эмоции, возбуждаемые музыкальными мелодиями, на возникновение синестезий? Ранее проведённые нами исследования показали, что около 50% испытуемых оценивали свой отклик на прослушивание мелодий с эмоциональным содержанием двухзначным ответом: или мелодия возбуждает, или успокаивает при прослушивании. Синестезии, являясь одним из способов расширения сферы восприятия человека, могут возникать при определённых условиях, соответствующих достаточно высокому уровню самоорганизации, гармоничному состоянию и эмоциональной поддержке. Поэтому для тестирования синестезий были привлечены испытуемые, имеющие обычно эмоциональный отклик на прослушивание музыкальных произведений. Каждому испытуемому предлагалось прослушать любимую мелодию, чтобы вызвать чёткую эмоциональную реакцию, наполненную уже знакомым и понятным содержанием.
При рассмотрении карт (рис.6. А В) привлекают внимание изменения в распределениях спектральной мощности на частотах альфа-ритма 7-8 Гц, 10-11 Гц и 11-12 Гц, которые возникают во время прослушивания музыкальной мелодии.
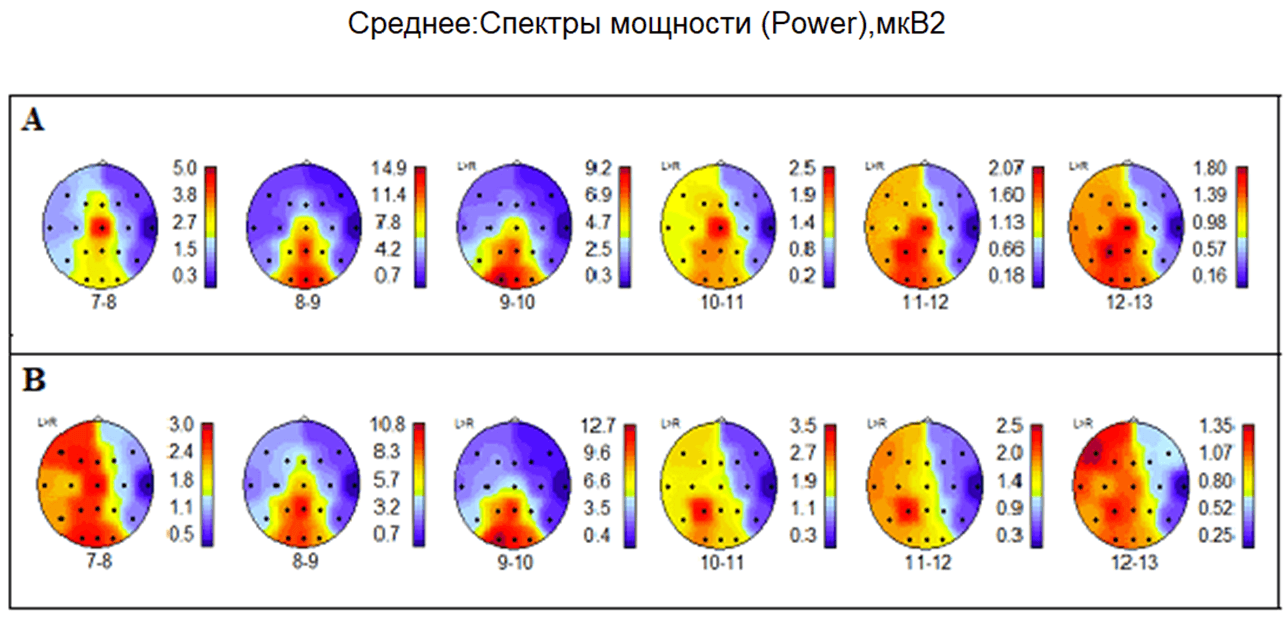
Рис. 6. Испытуемая М. Распределение спектральной мощности альфа-ритма. А – фон в состоянии спокойного бодрствования, психической и мышечной релаксации с закрытыми глазами. В – состояние во время прослушивания любимой музыкальной мелодии с закрытыми глазами. (Карты коры мозга ориентированы: лоб – сверху, затылок – снизу, справа – шкалы величин мкВ2, внизу – частота альфа-ритма в Гц). Подробности в тексте
Максимум мощности сенсомоторного альфа-ритма частотой 7-8 Гц ожидается в области центральных отведений, что и наблюдается в фоне в состоянии психической и мышечной релаксации с закрытыми глазами. Однако во время прослушивания любимой мелодии дополнительно возникают максимумы спектральной мощности в зрительных зонах затылочной коры, в центрах слухового восприятия височной доли, в ассоциативной теменной области. Можно предположить, что у испытуемой М. возможно возникновение синестезий телесно-зрительных, телесно-слуховых, зрительно-слуховых. Отмечаем также явное преобладание мощности альфа-ритма в левом полушарии не только на частоте 7-8 Гц, но и на частотах 10-11 Гц, 11-12 Гц и 12-13 Гц. Подобная межполушарная асимметрия обычно возникает при положительных эмоциях.
Для подтверждения развития синестезий во время прослушивания любимой музыкальной мелодии проведём сопоставление особенностей распределения спектральной мощности альфа-ритма выделенных частот с локализацией ЭДИ его происхождения и векторной направленностью.
На рис.7 (1,2,3) наблюдается межполушар-ная асимметрия распределения спектральной мощности альфа-ритма с преобладанием в левом полушарии. При рассмотрении локали-зации ЭДИ альфа-ритма на схемах мозга в декартовой системе координат (рис.7 1А, 2А, 3А) обнаруживается «парадокс» - основная концентрация ЭДИ в правом полушарии.
Первое предположение. Не исключено (рис.7.1А,2А,3А), что под влиянием эмоций, которые содержит мелодия, расширяется сфера восприятия и активируется правое полушарие для приёма и передачи более высокочастотных сигналов. Развивается реакция активации в виде десинхронизации, и амплитуда альфа-ритма снижается, падает мощность. Действительно, наибольшее падение мощности альфа-ритма наблюдается вследствие активации центрального слухового анализатора (поля Бродмана 41,42) в височной области Т4 отведения ЭЭГ. Это означает, что испытуемый находится в состоянии активного прослушивания музыкальной мелодии.
Второе предположение. Рис.7(1В, 2В, 3В) показывает, что вектора ЭДИ альфа-ритма направлены в сторону левого полушария. Если на частоте 7-8 Гц направленность векторов варьирует от затылка к теменным и лобно-височным областям, то на частотах 10-11-12 Гц вектора, в основном, направлены в теменную область противоположного полушария.
Известно [17,18], что характер потенциального поля при радиальном расположении диполя (один экстремум – максимум) зависит от глубины его расположения и определяет величину амплитуды на поверхности мозга в той области, куда направлены вектора данного диполя. На рис.7(1АВ, 2АВ, 3АВ) показан результат направленности векторов ЭДИ, локализованных преимущественно в области поясной извилины и медиальной поверхности височной доли (поля Бродмана 23,29,30,31,34,35,36,38) в правом полушарии на распределение спектральной мощности альфа-ритма в левом полушарии.
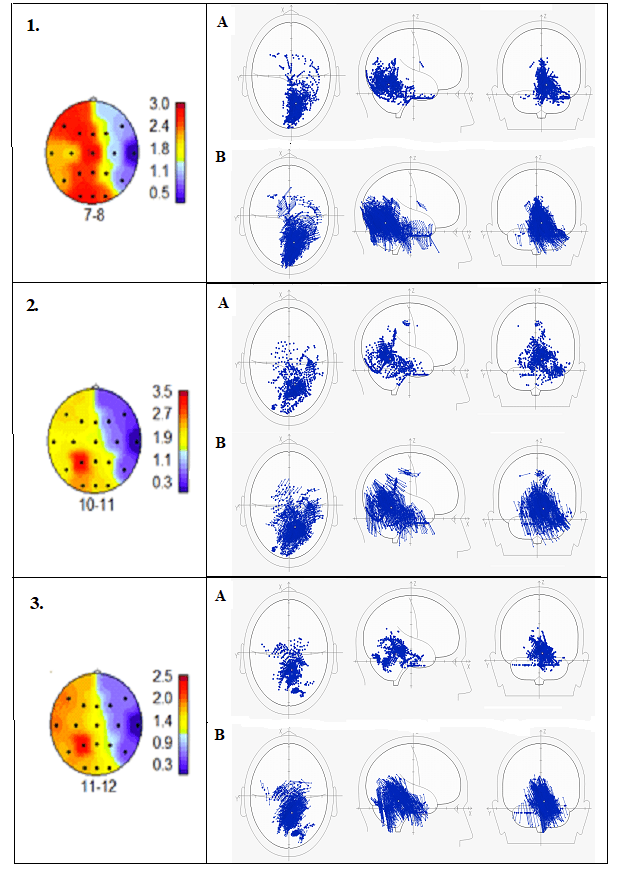
Рис. 7. Испытуемая М. 1,2,3 – нейрокартирование спектральной мощности альфа-ритма во время прослушивания любимой мелодии (карты взяты из рис.6.В для последующего анализа распределений ЭДИ и векторной направленности). А – на схематическом изображении мозга в декартовой системе координат представлено распределение эквивалентных дипольных источников (ЭДИ) происхождения альфа-ритма частотой, которая указана в п.п.1,2,3. В – те же ЭДИ, но с векторной направленностью. Подробности в тексте
Обратим внимание на то, что минимальная мощность альфа-ритма наблюдается в зоне слухового анализатора правой височной доли, так как здесь преобладает высокочастотная активность, связанная с эмоциональным восприятием любимой музыкальной мелодии. Подтверждено статистически достоверно [14], что в первичных зонах слухового восприятия (полях 41,42 по Бродману) преимущественно в правом полушарии регистрируются импульсы бета- и гамма-частоты ЭЭГ во время прослушивания музыкальных мелодий с эмоциональным содержанием. Колебания альфа-диапазона в этот период имеют несколько иные источники происхождения.
Так как в данной работе рассматриваются ЭДИ альфа-ритма с высоким уровнем дипольности КД=>0,98, то предполагается, что они с наибольшей вероятностью определяют более высокую активность отдельных структур мозга и их взаимодействие в альфа-диапазоне. ЭДИ альфа-ритма, согласно рис.7, локализованные в глубинных структурах правого полушария и вдоль центральной оси мозга, включая ядра восходящей неспецифической активации продолговатого мозга, срединных областей, отдельные ядра таламуса и центры эмоциональной активации в основании мозга, имеют электрические вектора, направленные в сторону коры мозга левого полушария, и определяют максимумы спектральной мощности на её поверхности. Чередование направленности диполей в альфа-ритме показывает, что электрические альфа-волны могут распространяться непосредственно в средах мозга, влияя на состояние возбудимости мембран отдельных элементов нейронных сетей [19]. Кроме того предполагается, что альфа-волны, обладая свойствами сканирования и стробирования [13], могут переносить информацию о возбуждении отдельных нейронных сетей из одной области мозга в другую, в частности в зоны ассоциативного восприятия, такие как поле 7 по Бродману или другие вторичные и третичные поля анализаторов. При рассмотрении синестезий было бы достаточно допустить, что взаимодействие нейронных сетей разных анализаторов первичного сенсорного восприятия обеспечивается анатомически с помощью синаптических контактов между аксонами, телами нейронов и дендритами. Однако возможна значительная потеря времени не только на распространение сигналов по нейронным путям, но и на передачу в синаптических контактах. На это обстоятельство первыми обратили внимание Мак Каллок и Винер [13, 20].
Известно, что распространение сенсорных сигналов от периферии к центру по афферентным анатомическим путям происходит унилатерально до уровня нейронов второго порядка, при этом аксоны нейронов второго порядка пересекают срединную линию, а затем направляются вверх и оканчиваются в ядрах таламуса контралатерального полушария. В рассматриваемом нами случае (рис.7) ЭДИ альфа-ритма с высоким уровнем дипольности локализуются в правом полушарии, то есть ипсилатерально по отношению к системе первичного слухового восприятия, если учитывать основные базовые функции полушарий головного мозга.
При высоком уровне дипольности размерность электрических полей ЭДИ достаточна, чтобы достичь поверхности коры противоположного полушария. Тем более отмечено, что если соотношения глубинных и поверхностных скальповых потенциалов Uглуб/Uсклп для альфа-ритма наиболее оптимальны и составляют только 1,2-1,5/1, то для дельта и тета-активности они составляют от 4/1 до 18/1 и даже до 80/1 [18]. По всей вероятности, эти тонические влияния в среде мозга не только обеспечивают повышение возбудимости ассоциативных путей для передачи информации из одной области в другую, но и способствуют формированию специфической активности с воссозданием внутренних образов и возникновением синестезий.
Синестезии возникают на уровне осознания. Нейрофизиологические исследования, проводимые с использованием объективных интегральных показателей активности мозга, позволяют изучить последовательность процессов, регулируемых сознанием, а также понять алгоритм деятельности мозга на подсознательном уровне. Можно распознать характер эмоциональных реакций по степени вовлечения в активность эмоциональных центров в базальных структурах мозга. Сопоставляя полученные данные с чувствами и ощущениями испытуемых во время прослушивания музыки и возникновения синестезий мы не находим противоречий, но осознаём, что это не просто совмещение образов разной модальности в пространстве взаимодействия нейронных сетей. Процесс творчества не ограничивается пространством индивидуального мозга. Для рождения новых истин требуются неповторимые ситуации.
Однако подобные представления возникают и у других исследователей синестезий. В качестве примера приводим следующее описание. «Подавляющее большинство синестетов-музыкантов «слышит» во время звучания произведения только один цвет – цвет тоники, центра акустических и функциональных тяготений. Следовательно, люди, обладающие цветным слухом, слышат в цвете не музыкальные высоты и не интервалику созвучий, а логическое и акустическое основание гармонической системы, в которой создано произведение, а именно его тонический аккорд. Синестетические ощущения у носителей цветного слуха исчезают при восприятии ими атональных или микрохроматических музыкальных произведений. Это явление закономерно в свете идеи первичной нерасчленимости перцепции и мышления. Гармония является грамматикой и логикой музыки, это главный уровень музыкального мышления. В случае цветного слуха смысл – это целостность гармонической системы и активность ее акустического центра» [9].
Выводы
- Достигнута основная цель работы, которая включает определение алгоритма деятельности мозга в процессе отдельных видов восприятия: слухового, зрительного, соматосенсорного, его особенности у лиц с доминирующим типом восприятия и при возникновении синестезий под влиянием эмоциональных музыкальных мелодий.
- В основе алгоритма лежит волновая функция альфа-ритма головного мозга и коммуникативные свойства этой функции.
- Особенностью альфа-ритма является специфичность его частоты для отдельных видов восприятия: зрительного 9…11 Гц и 11…13 Гц в первичной зоне зрительного восприятия, соматосенсорного 7…9 Гц в первичной зоне соматосенсорной чувствительности, слухового 7…9 Гц и 11…13 Гц в области центрального слухового анализатора. Эта специфичность имеет вероятностные границы для основных типов восприятия человека и является основой обмена информацией между зонами первичного восприятия.
- Вероятностные границы частотной специфичности альфа-ритма расширяются у лиц с доминирующим типом восприятия: у визуалов, аудиалов, кинестетиков и дискретов от 7 до 13 Гц в области центрального анализатора доминирующей сенсорной модальности.
- В качестве основных показателей волновой функции альфа-ритма рассмотрены распределения во времени или пути движения эквивалентных дипольных источников его происхождения и направления векторов отдельных колебаний.
- Сложный алгоритм, включающий частотную специфичность альфа-ритма, определяемую посредством спектрального анализа ЭЭГ, пространственную локализацию эквивалентных дипольных источников его происхождения, направленность их движения и векторов отдельных колебаний, был разработан как инструмент для изучения синестезий.
- Показано, что проявления синестезий помимо анатомически обеспеченного нейронного взаимодействия между зонами центральных анализаторов, ассоциативными центрами, структурами систем неспецифической восходящей активации и центрами эмоциональной активации могут быть обусловлены также непосредственными влияниями генерируемых в тканях мозга электрических токов и полей. Для изучения этих процессов были успешно использованы вычисления эквивалентных токовых диполей и распределения их электрических полей в качестве основных показателей альфа-ритма ЭЭГ.
- Эквивалентные дипольные источники происхождения колебаний альфа-ритма ЭЭГ с высоким коэффициентом дипольности (КД=>0,98) рассматриваются как наиболее активные локальные зоны, запускающие последовательное перемещение волновых процессов альфа колебаний. Эти колебания разности электрических потенциалов оказывают непосредственное влияние на организацию первичного восприятия в центральных анализаторах и могут сопровождать стробирование и перенос информации в ассоциативные зоны построения сложных образов.
- Основные объективные ЭЭГ показатели возможного возникновения синестезий определяются у каждого индивидуума в состоянии спокойного бодрствования с закрытыми глазами путём обнаружения в коре того или иного первичного сенсорного анализатора максимума мощности альфа-ритма той частоты, которая для него не специфична. Дополнительным показателем является векторная направленность эквивалентных токовых диполей альфа-ритма в сторону сенсорных анализаторов иной частотной специфичности.
- Прослушивание музыкальных мелодий с эмоциональным воздействием сопровождается индивидуальными изменениями максимумов спектральной мощности альфа-ритма в отдельных корковых центрах первичного сенсорного восприятия, распределений эквивалентных токовых диполей альфа-ритма и их векторной направленности, которые свидетельствуют о развитии синестезий.
