
Одной из важнейших задач нашего времени является сохранение биологического разнообразия лесов, так как оно представляет собой основу стабильности экосистем.
Наилучшие результаты для идентификации древесных видов дают анализ ядерных микросателлитных локусов и сравнение последовательностей фрагмента ITS ядерной рибосомной ДНК.
Целью данной работы является проведение идентификации видов и гибридов пихты, произрастающих в Ивантеевском дендрологическом парке им. акад. А.С. Яблокова.
Закладка Ивантеевского дендрологического парка им.акад. А.С. Яблокова началась в 1936 году. Деревья Abies holophylla и Abies veitchii были высажены в 1939 году, Abies sibirica в 1947, Abies sibirica x Abies veitchii в 1954, а Abies semenovii в 1960 году [1]. За прошедшие годы в посадках произошли выпады, не все записи сохранились, поэтому в таких случаях успешно проводят молекулярно-генетическую идентификацию.
Гербарий и материал для молекулярно-генетического анализа был собран в Ивантеевском дендрологическом парке им. академика А.С. Яблокова в октябре 2022 года с 17 деревьев, из которых предположительно 7 являлись гибридами Abies sibirica x Abies veitchii, 4 – принадлежали Abies holophylla, и по 2 дерева – Abies veitchii, Abies sibirica, Abies semenovii.
Для проведения микросателлитного анализа были отобраны три пары праймеров: SF324, SF333, SFb5 [2].
В результате проведения молекулярно-генетического анализа с тремя ядерными микросателлитными локусами и последующего генотипирования получена матрица с многолокусными генотипами для каждого из изученных образцов пихты.
Анализ генетических данных был проведен в программе GenAlEx v6.51. Аллельный состав каждого локуса приведен в таблице 1. Для локуса SF324 выявлено 3 аллеля, для локусов SF333 и SFb5 по 4 аллеля в каждом. Аллели, характеризующие тот или иной вид или гибрид, выделены жирным шрифтом.
Таблица 1
Размеры, аллельный состав и частоты аллелей трех ядерных микросателлитных локусов у образцов пихты
|
Локус |
Аллель и его размер, п.н. |
A. holophylla |
A. veitchii |
A. sibirica |
A. semenovii |
A. sibirica x A. veitchii |
|---|---|---|---|---|---|---|
|
SF324 |
107 |
1,000 |
0,000 |
0,000 |
0,000 |
0,000 |
|
113 |
0,000 |
0,500 |
1,000 |
1,000 |
1,000 | |
|
116 |
0,000 |
0,500 |
0,000 |
0,000 |
0,000 | |
|
SF333 |
152 |
0,000 |
0,000 |
0,500 |
0,000 |
0,100 |
|
154 |
0,500 |
0,250 |
0,000 |
1,000 |
0,000 | |
|
160 |
0,000 |
0,500 |
0,500 |
0,000 |
0,900 | |
|
162 |
0,500 |
0,250 |
0,000 |
0,000 |
0,000 | |
|
SFb5 |
120 |
0,000 |
0,500 |
0,000 |
1,000 |
0,000 |
|
126 |
0,000 |
0,000 |
1,000 |
0,000 |
0,500 | |
|
128 |
1,000 |
0,000 |
0,000 |
0,000 |
0,000 | |
|
130 |
0,000 |
0,500 |
0,000 |
0,000 |
0,500 |
На основе рассчитанных частот аллелей изученных трех ядерных микросателлитных локусов у образцов пихты, а также полученных генетических расстояний Нея выявлена степень сходства особей и их принадлежность к тому или иному таксону (таблица 2). Согласно генетическим расстояниям Нея, показана близость особей, определенных как A. veitchii и A. semenovii. Происхождение гибридов A. sibirica x A. veitchii и их родительские виды подтверждаются в данном анализе.
Таблица 2
Степень сходства изученных видов и гибридов пихты, выявленная на основе генетических расстояний Нея
|
A. holophylla |
A. veitchii |
A. sibirica |
A. semenovii |
A. sibirica x A. veitchii |
|
|---|---|---|---|---|---|
|
1,000 |
|
|
|
|
A. holophylla |
|
0,135 |
1,000 |
|
|
|
A. veitchii |
|
0,000 |
0,405 |
1,000 |
|
|
A. sibirica |
|
0,183 |
0,615 |
0,365 |
1,000 |
|
A. semenovii |
|
0,000 |
0,672 |
0,830 |
0,379 |
1,000 |
A. sibirica x A. veitchii |
Предварительные данные по изучению изменчивости трех ядерных микросателлитных локусов у образцов пихт показали, что в условиях дендропарка наиболее устойчивыми оказались вид A. veitchii и гибрид A. sibirica x A. veitchii, однако для подтверждения высокой биологической устойчивости данных таксонов необходимы дополнительные исследования в виде увеличения числа ядерных микросателлитных локусов.
Проверка генетических данных на выявление совпадающих генотипов показала их наличие. Все образцы, определенные как вид Abies holophylla, попали в одну группу. Вероятно, что посаженные рядом весной 1939 года саженцы пихты цельнолистной происходили из семян одного дерева, с одной или нескольких шишек, то есть эти образцы имеют общее, родственное происхождение и являются сибсами. В одну группу вошли все гибридные образцы. Образцы, определенные как Abies veitchii имеют различное происхождение.
В результате проведения молекулярно-генетического анализа на основе секвенирования фрагмента оперона ITS ядерной рибосомной ДНК получили сиквенсы длиной около 700 пар нуклеотидов для каждого образца. После выравнивания в программе BioEdit получили массив с последовательностями ДНК, который использовали для работы в программе MEGA для построения филогенетических деревьев с целью выявления сходства и родственных связей изученных образцов пихты. Филогенетические деревья строили различными способами (NJ, UPGMA, ME, MP, ML).
В качестве внешней группы из банка генетических данных взяли вид Abies concolor 1938002А [3].
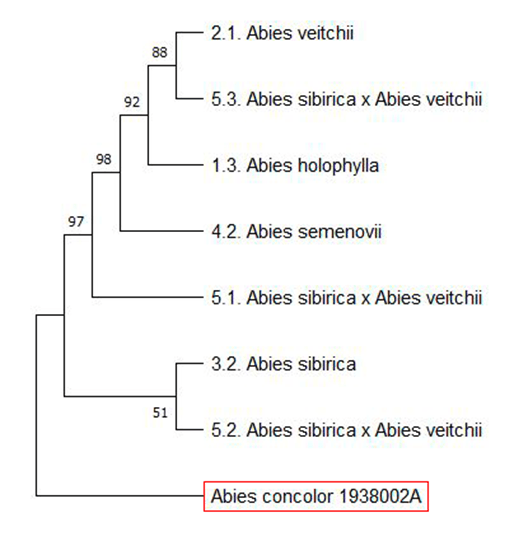
Рис. Дерево, построенное методом максимальной вероятности или максимального правдоподобия – Maximum Likelihood (ML). В узлах показаны значения бутстрэп-поддержки
С лучшей разрешающей способностью оказались деревья, построенные методами NJ – neighbor joining (ближайшего соседа) и ML – maximum likelihood (максимального правдоподобия) (рис.).
На основе генотипирования трех микросателлитных локусов ядерного генома образцов пихты Ивантеевского дендропарка составлены генетические паспорта.
Данные по молеулярной идентификации изученных видов и гибридов пихт будут переданы руководству Ивантеевского дендропарка.
